Joint research between Kobe University and National Hospital Organization Kyushu Cancer Center has revealed that mice with mutations in the YAP signal pathway develop head-and-neck cancer over an extremely short period of time (world’s fastest cancer onset mouse model), indicating that this pathway plays a crucial role in the onset of these cancers. This discovery may shed light on the development of new drugs for head-and-neck cancer.
This research resulted from a collaboration between a research group led by Professor SUZUKI Akira and Associate Professor MAEHAMA Tomohiko at Kobe University Graduate School of Medicine, and Dr. MASUDA Muneyuki’s team at Kyushu Cancer Center.
These results were published in the American scientific journal ‘Science Advances’ on March 18.
Main points
- Deletion of MOB1 (*1, which represses YAP) in mouse tongues causes strong activation of YAP (*2), leading to the early onset of cancer (in about 1 week).
- In humans, the expression of YAP increases during the development of dysplasia (pre-cancerous lesions), prior to the onset of head-and-neck cancer. YAP continues to increase with the development and progression of cancer. This high YAP activation is linked to poor patient prognosis.
- The onset and progression of head-and-neck cancer in the mice in this study, and the proliferation of stem cells in this cancer in humans, are dependent on YAP.
- These results suggest that cancer develops when the YAP activation exceeds a threshold. YAP may play a fundamental role in head-and-neck cancer onset and progression. These conclusions represent a paradigm shift in the understanding of these cancers.
- The mouse model developed in this study can be used in research to develop new drugs for head-and-neck cancer and, in addition, provides a beneficial resource for cancer research in general.
- By inhibiting YAP, the development and progression of head-and-neck cancer can be suppressed. Thus, the YAP pathway provides a good target for head-and-neck cancer treatments.
Research Background
Head-and-neck cancer in humans
Head-and-neck cancer is the sixth most common type of cancer in the world, affecting 600,000 people annually. In Japan there are around 22,500 new cases every year. This ‘head and neck’ includes the oral cavity and areas of the throat (pharynx and larynx). Among these, mouth cancers (especially tongue cancer) are the most prevalent.
It is understood that exposure to carcinogens, such as those found in cigarettes and alcohol, as well as mechanical irritation of the mucous membranes in the mouth, tooth decay and improperly fitted dentures, are risk factors for the development of head-and-neck cancer.
In addition, 15% of head-and-neck cancer is caused by Human Papillomavirus (HPV), which in particular causes oropharynx cancer.
The prognosis for patients who are HPV-positive is relatively good. Conversely, prognosis is poor for HPV negative patients and in most cases, mutations are found in the tumor suppressor gene TP53 (p53). However, mutations in this gene alone are not sufficient to cause head-and-neck cancer. It has been thought that changes in other molecules are also necessary for cancer development, however these causes remain elusive.
From comprehensive cancer genome analyses, it is known that PTEN/P13K (46%), FAT1 (32%), EGFR (15%) gene mutations are also found in HPV-negative head-and-neck cancer. However, the genetic pathway of these molecules in relation to head-and-neck cancer development has not been sufficiently understood.
Mouse models of cancer
Up until now, research using mouse models of head-and-neck cancer has discovered that if both the p53 and Akt genes are mutated, 50% of mice will develop this type of cancer about 9 months after the mutation (the average mouse lifespan is 2 years).
The onset of cancer begins after many genetic mutations have accumulated (multistep carcinogenesis). Mice with a mutation in one important molecule usually develop cancer within 4 to 24 months (with the majority showing signs between 6 to 12 months).
The YAP pathway
The function of the transcriptional co-activator YAP is to turn ‘on’ the transcription of gene clusters related to cell growth. The LATS/MOB1 complex phosphorylates YAP, thereby excluding YAP from the nucleus, leading to the subsequent degradation of YAP proteins. In other words, MOB1 and LATS act as a ‘brake’ (tumor suppressor) to inhibit cell proliferation facilitated by YAP. It has been reported that in 8% of human head-and-neck cancer cases, the YAP gene is amplified and there is a connection between YAP activation, cancer progression and poor prognosis.
This research group produced mice with MOB1 deletion in their tongues (so that YAP would be intrinsically activated) in order to perform a detailed analysis in vivo of the role that the YAP pathway plays in head-and-neck cancer.
Research Methodology
Mice with MOB1 deletion exhibit rapid onset tongue cancer
This research group developed mice with MOB1 deletion in their tongues by applying the drug tamoxifen to their tongues and then modifying them genetically using the Cre-loxP system (*4).
Three days after applying tamoxifen, the amount of MOB1 had barely decreased, however by day 7, the vast majority of these proteins had disappeared. At this point, a third of the mice demonstrated rapid onset head-and-neck cancer (intraepithelial tongue cancer), with all mice developing the disease by day 14. The cancer had progressed in all mice by day 28 (invasive tongue cancer). The team succeeded in developing the world’s fastest mouse model of cancer onset (Figure 1). Both domestic and international patents for this model have been applied for.
This mouse model showed that head-and-neck cancer develops quickly (within a week) when the YAP pathway is strongly activated, suggesting that this pathway plays an extremely important role in head-and-neck cancer onset.
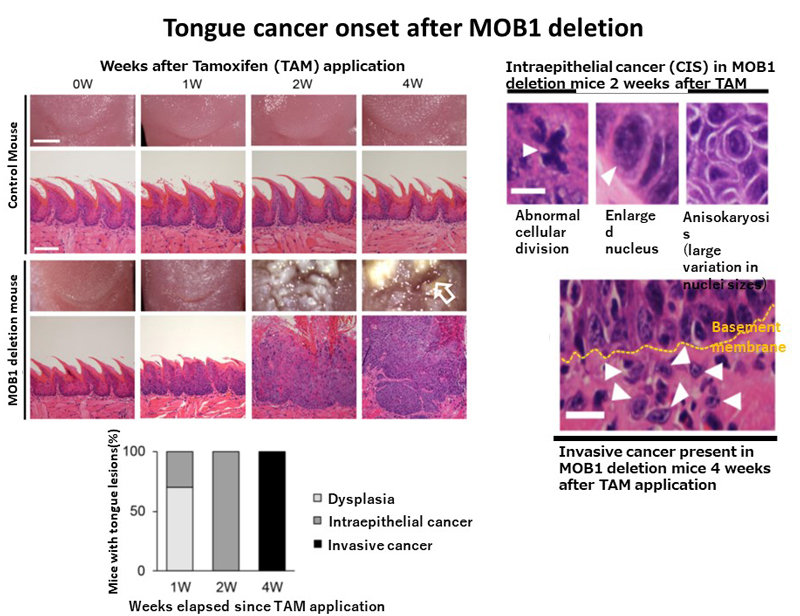
This research group developed mice with MOB1 deletions in their tongues through Tamoxifen application. Seven days after the drug application, approximately a third of the mice developed early onset head-and-neck cancer (intraepithelial tongue cancer), with all mice developing it by day 14. By day 28, all mice had invasive tongue cancer, making this the world’s fastest model of cancer development in mice.
YAP activation and tumorigenic properties of the tongue epithelium in MOB1 deletion mice.
The epithelial cells (on the surface of the tongues) of MOB1 deletion mice exhibited the following properties characteristic of tumor development: increased cell proliferation and cell saturation density, impaired cell polarity, low levels of apoptosis (cell death), increase in undifferentiated cells, and chromosomal instability (characterized by increases in aneuploid cells (*5)), multipolar spindles (*6) and micronucleated cells). On a biochemical level, activation of YAP and a decrease in LATS proteins was evident due to MOB1 deletion.
The epithelial cells acquired the characteristics of tumor cells due to the YAP activation caused by the deletion of MOB1.
YAP activation in the stages of tongue cancer in humans
The development of human tongue cancer can be divided to the following stages; the normal stage, the dysplasia stage, the intraepithelial cancer stage (*8) and the invasive cancer stage (*9).
If we look at YAP activation across all these stages, we can see that YAP is enhanced in the dysplasia stage which proceeds the onset of cancer. YAP activation shows continued increase during the subsequent stages of cancer progression. In cases where YAP is highly activated, overall survival is decreased and the likelihood of cancer relapse is high.
In other words, YAP increases before the onset of cancer and continues to increase as the cancer develops and progresses. Accumulation of YAP is linked to poor patient prognosis.
Cancer formation is dependent on YAP when MOB1 is deleted
Invasive cancer occurred in MOB1 deletion mice. However, when both YAP and MOB1 are deleted from mice, cancer onset is halted at the dysplasia stage, showing that the onset of head-and-neck cancer is dependent on YAP (Figure 2).
Among current YAP pathway inhibitors, the SRC inhibitor Dasatinib (*10) was shown to be the most effective (SRC has been previously shown to activate YAP both directly and indirectly). Dasatinib was shown to prevent the onset of intraepithelial head-and-neck cancer in the MOB1 deletion mice. It also suppressed the development of invasive cancer in MOB1 deletion mice that had reached the intraepithelial head-and-neck cancer stage.
In human head-and-neck cancer stem cells, it is possible to suppress cell proliferation either by inhibiting YAP gene expression or by adding YAP inhibitors. Cisplatin, which is commonly used to treat head-and-neck cancer, is augmented when YAP is suppressed.
In mice, head-and-neck cancer onset and progression was suppressed when YAP was inhibited. In the same way, it was shown that in human tongue cancer stem cells, cell proliferation was also suppressed when YAP was inhibited.
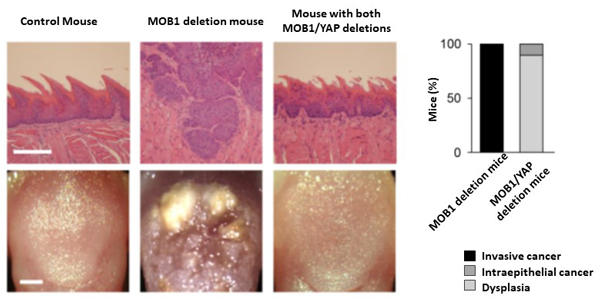
Invasive cancer development was seen in MOB1 deletion mice. However, when both MOB1 and YAP are deleted, mice only develop dysplasia. This shows that cancer development is dependent on YAP.
Known genetic mutations in human head-and-neck cancer and YAP activation
Genetic mutations in p53, PTEN/PI3K, FAT1, and EGFR have been identified in HPV-negative head-and-neck cancer.
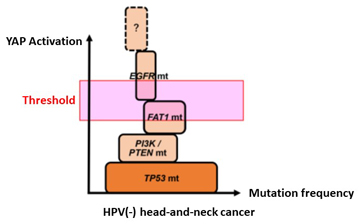
The head-and-neck cancer onset/progression process: genetic mutations in p53, PTEN/PI3K, FAT1 and EGFR accumulate causing YAP activation to reach the threshold for the likely onset of cancer.
This research group showed that EGF signal activation and mutations in p53, PTEN and FAT1 each play a role in YAP activation. Furthermore, YAP activation gradually increases as these genetic mutations accumulate.
Normally, cancer takes time to develop as it is a multistep process. However, in this study, intraepithelial head-and-neck cancer rapidly developed just from highly strengthening YAP activation.
In conclusion, this study raises the possibility that the following process for head-and-neck cancer development takes place: A. Cancer develops when the YAP activation exceeds a threshold due to the accumulation of genetic mutations in p53, PTEN/PI3K, FAT1 and EGFR (Figure 3). B. Subsequently, YAP continues to accumulate after cancer has developed, resulting in cancer progression.
Conclusion and Further Developments
YAP is frequently activated in cancer cells although genetic mutations in the YAP pathway are not frequently found. It is thought that this is why the importance of the YAP pathway in the onset of head-and-neck cancer was unclear until now.
1. YAP activation levels are high before the onset of head-and-neck cancer in humans.
2. YAP is further activated as the cancer progresses.
3. The high frequency of mutations in p53, PTEN/PI3K, FAT1 and EGFR all activate YAP.
4. The accumulation of these molecular mutations gradually leads to high YAP activation:
A. The accumulation of genetic mutations in p53, PTEN/PI3K, FAT1 and EGFR cause YAP to reach its threshold, culminating the onset of cancer.
B. YAP continues to accumulate after the cancer onset, resulting in further cancer progression.
It is necessary to consider YAP as a basis for head-and-neck cancer onset and progression. This represents a paradigm shift in our understanding of these cancers.
In addition, it has also been shown that risk factors for head-and-neck cancer, such as cigarette smoking, mechanical irritation of mucous membranes and HPV infection, also play a part in YAP activation.
The mouse model in this study: 1. Is the fastest mouse model in the world for showing the natural onset of cancer. 2. Can be used to visualize cancer onset and progression. 3. Allows cancers to be developed naturally at the same time. 4. Allows cancer onset and progression to be analyzed in mice immediately after birth, allowing drug tests to be conducted in a shorter period of time and in small quantities. The results suggest that this mouse model would be ideal, not only for research into developing new treatments for head-and-neck cancer, but also for cancer research in general.
It is expected that the YAP pathway will provide a good target for drugs used in the treatment of head-and-neck cancer because inhibiting YAP not only suppresses the cancer onset but can also prevent its progression.
Researchers from all over the world, including this research group, are currently trying to find new drugs that target the YAP pathway. We have shown one factor that is effective against head-and-neck cancer. It is also expected that the mouse model will become an indispensable tool for evaluating their results and for head-and-neck cancer research.
Glossary
- 1. MOB1 (Mps One Binder 1)
- MOB1 is necessary for activating the LATS kinase and acts as a brake (tumor suppressor) on downstream, negatively-controlled YAP. Deletion of MOB1 exacerbates cell proliferation and leads to cancer.
- 2. YAP (Yes-associated Protein)
- YAP is a transcriptional coactivator. It forms a complex with multiple transcription factors inside the nucleus to control the expression of various genes. YAP is phosphorylated by the LATS kinase, causing YAP to be excluded from the nucleus and inactivated.
- 3. Human Papillomavirus (HPV)
- A type of papillomavirus, there are over a hundred genotypes or varieties of HPV. The virus is linked to genital warts, head-and-neck cancer and cervical cancer.
- 4. Cre-loxP system
- A genetic modification system using Cre recombinase, which catalyze DNA recombination between two loxP sites (a 34-base pair nucleotide sequence). If Cre is expressed in a cell where chromosomal DNA has been artificially inserted into two loxP sites, the intervening DNA segment is deleted.
- 5. Aneuploid Cells
- contain an abnormal number of individual chromosomes. This does not include a difference of one or more complete sets of chromosomes.
- 6. Multipolar Spindle
- Spindles are formed during cell division to separate chromosomes between daughter cells. In normal cells, a pair of centrosomes forms prior to cell division to organize proteins called microtubules into a spindle between the two centrosomes. However, in cancer cells there are more than two centrosomes, and so-called multipolar spindles form. This can cause chromosomal instability and lead to the formation of aneuploid cells because the chromosomes were not correctly allocated at the time of cell division.
- 7. Dysplasia
- The presence of cells of an abnormal type within a tissue. In medical diagnosis, the presence of abnormal-looking cells (dysplasia) observed under a microscope can signify an increased chance of a patient developing cancer (pre-cancerous symptoms).
- 8. Intraepithelial cancer
- This is where cancer cells are found within the intraepithelial layer of cells which form on an organ’s surface. At this point, cancer cells have not been able to penetrate the basement membrane and have not spread deeply. Related terms include Carcinoma in Situ (CIS), intraepithelial tumor and intraepithelial neoplasia.
- 9. Invasive cancer
- is where cancer cells penetrate the basement membrane, a thin membrane separating them from other tissues. From there, cancer can spread to the surrounding area.
- 10. Dasatinib
- a chemotherapy drug that inhibits the SRC kinase family, which can cause malignant transformation in cells. SRC is both directly and indirectly connected to YAP activation, so dasatinib can also inhibit YAP. It is currently used to treat leukemia but is not prescribed for head-and-neck cancer treatment.
Acknowledgements
This research was made possible primarily through funding to the project ‘The development of cancer treatment methods targeting cancer suppression genes.’ (Lead researcher: Suzuki Akira) as part of the Japanese Agency for Medical Research and Development’s Project for Cancer Research and Therapeutic Evolution (AMED P-CREATE).
Journal Information
- Title
- “YAP1 is a Potent Driver of the Onset and Progression of Oral Squamous Cell Carcinoma”
- DOI
- 10.1126/sciadv.aay3324
- Authors
- Hirofumi Omori, Miki Nishio, Muneyuki Masuda, Yosuke Miyachi, Fumihito Ueda, Takafumi Nakano, Kuniaki Sato, Koshi Mimori, Kenichi Taguchi, Hiroki Hikasa, Hiroshi Nishina, Hironori Tashiro, Tohru Kiyono, Tak Wah Mak, Kazuwa Nakao, Takashi Nakagawa, Tomohiko Maehama*, Akira Suzuki*
(*Corresponding authors) - Journal
- Science Advances