理化学研究所 (理研) 生命機能科学研究センター形態進化研究チームの樋口真之輔研修生 (神戸大学大学院理学研究科大学院生)、倉谷滋チームリーダー (開拓研究本部倉谷形態進化研究室主任研究員)、兵庫医科大学の菅原文昭講師、東京大学大気海洋研究所の高木亙助教らの共同研究グループ※は、顎 (あご) を持たない脊椎動物であるヌタウナギ[1] とヤツメウナギ[2] の内耳の発生を解析し、単一の半規管[3] から段階的に複雑に進化したと考えられてきた三つの半規管 ( 三半規管[3] ) の構成要素の大部分が、すでに5億年以上前の共通祖先において獲得されていたことを明らかにしました。
本研究成果は、ヒトの聴覚・平衡感覚を担う内耳の進化について理解を深めるとともに、感覚器の発生生物学研究にも貢献すると期待できます。
今回、共同研究グループは、ヒトと5億年以上前に分岐した、顎を持たない脊椎動物であるヌタウナギとヤツメウナギにおける内耳の形成過程を詳細に観察し、前半規管と後半規管の位置を決める遺伝子の発現が、両動物でよく似たパターンを示すことを見いだしました。さらに、両動物胚には三つ目の半規管である水平半規管がまだないものの、そのセンサーとなる感覚上皮[4]に相当する構造がすでに存在することを確認しました。これにより、脊椎動物の共通祖先が、前後二つの半規管を形成する発生プログラムをすでに持っており、さらに既存の感覚上皮を転用して三つ目の水平半規管とその検出器を獲得し、3次元方向の回転に対応した三半規管へと進化したことが明らかになりました。
本研究は、英国の科学雑誌『Nature』の掲載に先立ち、オンライン版 (12月5日付け:日本時間12月6日) に掲載されました。
背景
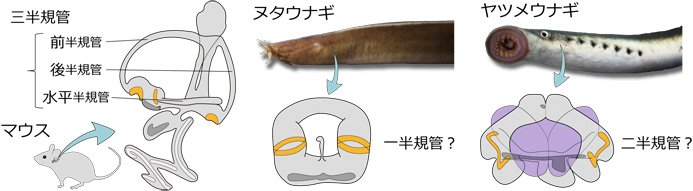
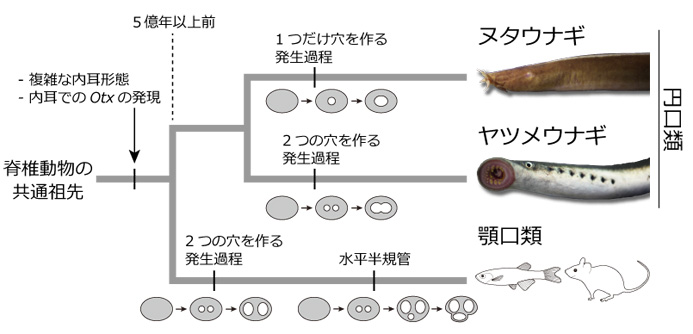
動物は、光、音、化学物質など身の回りの情報を得るために、目、耳、鼻といった感覚器を進化させてきました。これらの中で耳 (内耳) は、ヒトでは聴覚器としても働きますが、脊椎動物の進化の歴史においては、重力や頭部の回転運動を感知する平衡感覚器として出現しました。ヒトやカエル、サメなど顎 (あご) を持つ脊椎動物 (顎口類) の内耳には、平衡感覚を担う三半規管があります。三半規管はその名のとおり、三つの半円状の管 (半規管) が互いに直交し、それぞれに検出器である感覚上皮が一つずつ付属し、三次元方向の回転を感知しています。これに対し、顎を持たない脊椎動物であるヌタウナギは一つの半規管、ヤツメウナギは二つの半規管しか持たず、古典的には半規管の数が1→2→3と段階的に進化したと解釈されてきました (図1左)。しかし近年、ヌタウナギとヤツメウナギが「円口類」(顎を持たない現生の脊椎動物) という一つのグループに属し、どちらかがより「古い」とはいえないことが分かり (図1右)、それとともに三半規管の進化過程は再び謎に包まれることになりました。
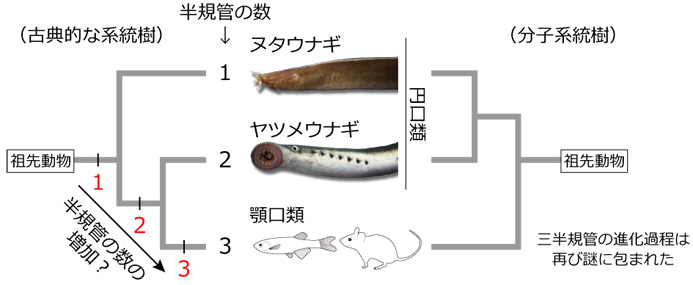
古典的には、左側の系統樹で示すように、ヌタウナギは他の脊椎動物よりも「古い」グループであり、半規管の数は1→2→3 (赤) と進化したと考えられてきた。ところが、遺伝子配列の比較により、ヌタウナギとヤツメウナギは「円口類」という一つのグループであることがわかった。つまり、右側の系統樹で示すように、単純な構造に見えるヌタウナギの一つの半規管は、必ずしも「祖先的だ」とはいえない。
研究手法と成果
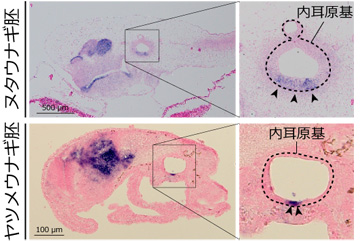
内耳の前後方向を決める遺伝子 (Tbx1/10A) はヌタウナギ、ヤツメウナギの両方で内耳原基 (点線) の後方に発現する (右側の青い染色の部分)。写真の左方向が前側 (頭部側)。
今回、共同研究グループは、過去に考えられてきたような、半規管の数が1→2→3と増加する上昇的な進化ではなく、5億年以上前に存在した脊椎動物の共通祖先が、すでに複雑な構造の内耳を持っていたことを明らかにしました。
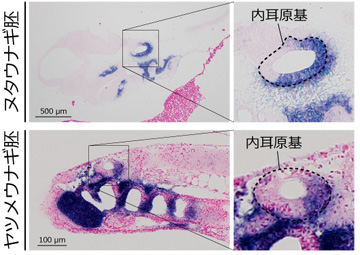
まず、ヌタウナギとヤツメウナギの胚において、内耳の発生過程を詳細に観察したところ、顎口類において前半規管と後半規管の位置を決める遺伝子が、円口類の2種でもよく似たパターンで発現していることが分かりました (図2)。つまり、すべての脊椎動物の共通祖先は、もともと前後に分かれた二つの半規管を「作りうる」発生プログラムを持っていたわけです。
ただし、実際に前後二つの半規管ができるためには、内耳の2カ所に穴が開き、二つの環が形成される必要があります。本研究により、ヌタウナギでは内耳原基[5]の中央に一つだけ穴が貫通し、一つの環状の半規管が直接生じることが初めて明らかになりました。これに対して、ヤツメウナギでは顎口類と同じく、分離した二つの半規管が一時的に現れることが分かりました。これらの比較から、ヌタウナギの単純な発生過程は、ヌタウナギが独自に獲得したものと推定できます (図3)。
5億年以上前に存在した脊椎動物の共通祖先は、すでに二つの半規管を作りうる発生プログラムと三つの感覚上皮を含む、複雑な内耳を持っていたことが分わかった。ヌタウナギ、ヤツメウナギの内耳の腹側部でOtx遺伝子発現が認められたことから、顎口類の水平半規管は、Otx遺伝子の新たな発現によってではなく、Otxを必要条件とする発生過程の変化により獲得されたと考えられる (図4)。
顎口類の三半規管のうち、水平半規管の発生に必要な遺伝子Otx1の相同遺伝子OtxAは、ヌタウナギ、ヤツメウナギの両方で内耳原基の腹側部に発現する (右側の点線で囲っている部分が内耳原基、黒矢頭で示した青い染色の部分がOtxAの発現示す)。この発現部位は、顎口類では水平半規管が形成される領域に対応すると考えられる。
次に、顎口類が持つ第3の半規管、水平半規管の進化に着目しました。水平半規管は、顎口類の発生過程では内耳の腹側部に形成されます。転写因子[6]の一つをコードするOtx1遺伝子のノックアウトマウス[7]では水平半規管のみが欠失することから、Otx1が水平半規管の発生に必要だと考えられてきました注1)。
そこで、円口類でOtx1の相同遺伝子[8]の一つであるOtxAの発現を観察すると、驚いたことにヌタウナギとヤツメウナギの両方の内耳の腹側部でOtxAが発現していました (図4)。つまり、顎口類における水平半規管の獲得は、Otx1が内耳の腹側部に新たに発現するようになったからではなく、もともとここに発現していたOtx1を必要条件として、内耳の発生過程が変化し成立したためと考えられます。この仮説については、今後さらなる解析により検証が必要です。
注1) Fritzsch, B., Signore, M. & Simeone, A. Otx1 null mutant mice show partial segregation of sensory epithelia comparable to lamprey ears. Dev. Genes Evol. 211, 388–396 (2001).
最後に、顎口類の水平半規管に付属する感覚上皮に着目しました。従来から、ヤツメウナギには「背側斑」と呼ばれる感覚上皮とそれを中枢に伝える神経を持つことが知られていましたが、今回、これらが顎口類の水平半規管の感覚上皮と極めて似た発生過程を経ることが明らかになりました。さらに、ヌタウナギでも同様の感覚上皮と神経枝を見つけたことから、円口類は水平半規管の環こそ完全に欠くものの、その検出部となる感覚上皮をすでに備えていたことが分かりました。
以上のことから、脊椎動物の共通祖先は、二つの半規管を作りうる発生的プログラムと、三つの半規管に対応する感覚上皮がすでに用意されており、顎口類ではこれらを巧みに利用して、三次元方向に対応した三つの半規管と感覚上皮を作り上げたことが分かりました。脊椎動物の共通祖先における内耳は、従来の説よりもはるかに複雑であったことが判明し、本研究では半規管の進化過程の一端を明らかにすることができました。
今後の期待
今回の研究により、内耳はこれまで考えられてきたよりも複雑な進化の歴史を辿って現在の形が成立したことが分かりました。すなわち、ヒトがもつような複雑な三半規管の形態を作り出す材料の起源は、円口類と顎口類の分岐以前の段階である5億年以上前にまで遡ることになります。しかしながら、一見単純な構造に見える円口類の内耳とヒトが持つ複雑な内耳には、どのような発生プログラムの違いがあり、おのおのがいかに進化したかについては未解決の問題です。
内耳の構造は化石として残りやすく、古脊椎動物学において、化石魚類の同定や進化過程を推定する上での鍵となる形質として着目されます。脊椎動物の進化の初期に現れた化石魚類の内耳を本研究で得られた理解に基づいて研究することで、これまで未分類であった化石動物の研究に貢献すると期待できます。
補足説明
- [1] ヌタウナギ
- 顎を持たない脊椎動物の一群。細長い体型から「ウナギ」と呼ばれるが、真骨魚ウナギ目の仲間ではない。明瞭な脊椎骨や眼のレンズを欠くため、かつてはヤツメウナギや他の脊椎動物よりも祖先的な動物だと考えられてきたが、現在はヤツメウナギとともに「円口類 (えんこうるい) 」という一つのグループをなすことが分かっている。
- [2] ヤツメウナギ
- ヌタウナギとともに円口類に属する一群。眼の後ろにある7対のえら孔を眼に見立ててこう呼ばれる。
- [3] 半規管、三半規管
- 脊椎動物の内耳に存在する平衡感覚器の一部。現生の顎口類では前半規管、後半規管、水平半規管の三つがある。この三つの半規管を合わせて三半規管と呼ぶ。
- [4] 感覚上皮
- 外部からの刺激を受容し、神経系へ伝達する上皮細胞。内耳の半規管では、有毛細胞が回転運動を感じるセンサーの役割を持つ。他には網膜、嗅上皮などが感覚上皮に該当する。
- [5] 内耳原基
- 発生の過程で、頭部の上皮にある特定の領域 (耳プラコード) が陥入してできたボール状の構造。耳胞とも呼ばれる。これがさらに形を変え、複雑な形の内耳ができる。
- [6] 転写因子
- DNAの特定の配列に結合するタンパク質。他の遺伝子の発現を促進または抑制する。
- [7] ノックアウトマウス
- 人為的に特定の遺伝子を削除したマウス。注目する遺伝子の働きを研究するために利用される。
- [8] 相同遺伝子
- 同一の祖先に由来する遺伝子のこと。
共同研究グループ
- 理化学研究所
生命機能科学研究センター 形態進化研究チーム- チームリーダー 倉谷 滋 (くらたに しげる)
(開拓研究本部 倉谷形態進化研究室 主任研究員) - 研修生 樋口 真之輔 (ひぐち しんのすけ)
(神戸大学大学院 理学研究科生物学専攻 大学院生)
開拓研究本部 倉谷形態進化研究室 - 研究員 Juan Pascual-Anaya (パスクアル アナヤ フアン)
脳神経科学研究センター 触知覚生理学研究チーム - 基礎科学特別研究員 大石 康博 (おおいし やすひろ)
- チームリーダー 倉谷 滋 (くらたに しげる)
- 兵庫医科大学 生物学
- 講師 菅原 文昭 (すがはら ふみあき)
(理化学研究所 開拓研究本部 倉谷形態進化研究室 客員研究員)
- 講師 菅原 文昭 (すがはら ふみあき)
- 東京大学 大気海洋研究所 海洋生命科学部門 (生理学分野)
- 助教 高木 亙 (たかぎ わたる)
研究支援
本研究は、日本学術振興会(JSPS)新学術領域研究「進化の制約と方向性 ~微生物から多細胞生物までを貫く表現型進化原理の解明~ (領域代表者:倉谷滋)」による支援を受けて行われました。
論文情報
- タイトル
- “Inner ear development in cyclostomes and evolution of the vertebrate semicircular canals”
- DOI
- 10.1038/s41586-018-0782-y
- 著者
- Shinnosuke Higuchi, Fumiaki Sugahara, Juan Pascual-Anaya, Wataru Takagi, Yasuhiro Oisi and Shigeru Kuratani
- 掲載誌
- Nature