神戸大学大学院農学研究科の三宅親弘教授、和田慎也助教、門田かなえ修士課程修了生と、東北大学大学院農学研究科の牧野周教授、岩手大学農学部の鈴木雄二准教授らの研究グループは、植物が活性酸素 (ROS) を抑制し酸化障害を防ぐ「P700酸化システム」において、重要な要素であるP700酸化で誘導される循環的電子伝達反応 (CEF) が、光化学系I (PSI) 複合体内部で生じる電荷再結合反応であることを解明しました。またさらに、電荷再結合反応によるROS生成抑制メカニズムも併せて明らかにしました。
これは、光合成電子伝達反応制御に関する研究の歴史の中で、これまで解明されていなかった生理現象です。
本研究成果は、6月5日に、科学誌「plants」に掲載されました。
ポイント
- 光合成は、酸化障害をもたらす活性酸素生成の危険性を含んでいる。
- 三宅親弘教授を代表とする研究チームはこれまでの研究で、植物が酸化障害を防ぐ「P700酸化システム」と呼ばれるメカニズムを持っていることを明らかにした。
- 本研究では、P700酸化に伴う光化学系I (PSI) での循環的電子伝達反応 (CEF) の誘導が、電荷再結合反応であることを明らかにした。
- 本研究における二つ目の成果として、電荷再結合反応によるROS生成抑制メカニズムを解明した。
研究の背景とこれまでの経緯
光合成が行われる時、太陽光のエネルギーは、CO2から糖を合成するためにチラコイド膜 ※1の光合成電子伝達系で化学エネルギー (NADPH, ATP) へ変換されます。この光合成電子伝達系には、光化学系I (PSI) およびII (PSII) があり、それぞれが光エネルギーを吸収しています。
PSIIでは、水を光酸化し、電子を引き抜くとともにO2を発生させています。PSIでは、PSIIで水から引き抜かれた電子を用いてNADP+をNADPHへ還元するために、フェレドキシン (Fd) ※2へ電子を渡す反応が進行しています。これらの水からNADPHへの電子の流れはリニア電子伝達反応と呼ばれ、光合成生物にとっては炭素獲得のための必要不可欠な反応です。
ところがこの反応は、光合成生物にとって酸化障害を起こしかねない危険な反応でもあります。1951年にPSIでの活性酸素 (ROS) 生成が発見され、以来、PSIで生成されるO2の危険性が議論されてきました。過去の議論では、光合成の進行が停滞する状況 (光合成生物への環境ストレス:低温、高温、乾燥など) では、PSIでNADPHが使用されず、電子が蓄積し、蓄積した電子がO2を還元した結果、スーパーオキシドラジカル (O2-) の生成をもたらすと考えられてきました。O2-は、ROSによる酸化障害のトリガー分子であり、さらなる高い反応をもつ過酸化水素H2O2、ヒドロキシルラジカルの生成にいたる要因となります。
三宅親弘教授を代表とする研究チームはこれまでの研究で、全てのO2発生型光合成生物がこの酸化障害を抑制する分子メカニズム「P700酸化システム」を持つことを明らかにしてきました。P700酸化という生理現象は、80年代の終わりから報告がありましたが、そのメカニズムや役割の解明に成功したのはこの研究チームが世界で初めてでした。
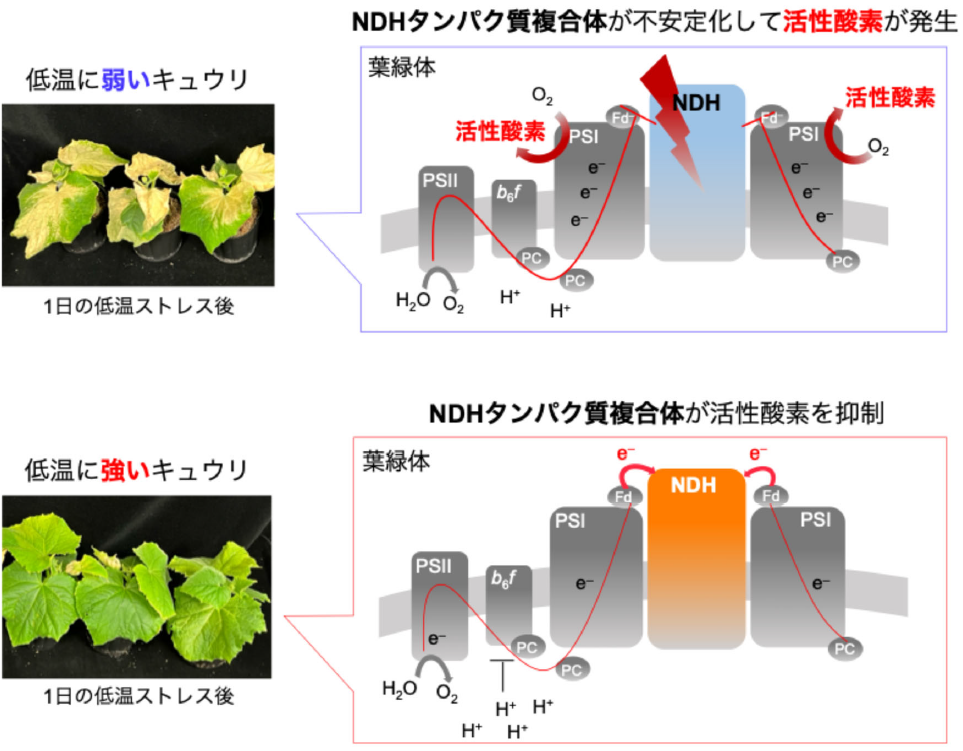
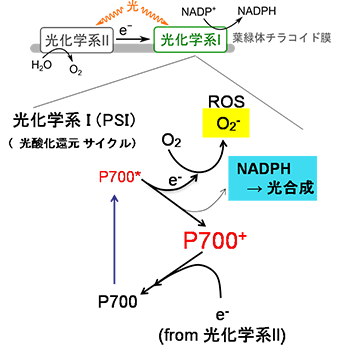
研究チームはまず、酸化障害を人工的に生じさせる方法(パルス法)を開発し、パルス法を用いて活性酸素 (ROS) 生成メカニズムを解明しました (図1)。光合成が抑制された場合、電子がNADP+へ流れずP700* が蓄積し、その結果、ROSであるスーパーオキシドラジカル (O2-) が過剰に生成されます。
そして次に、ROS生成を抑制する「P700酸化システム」とそのメカニズムも明らかにしました。PSIの反応中心クロロフィル ※3 であるP700は、光酸化還元サイクルで機能しています。光酸化還元サイクルでは、太陽光エネルギーがP700へ渡されることで、P700* (励起型P700) が生成され、P700*は電荷分離してNADP+に電子を渡し、自身はP700+ (酸化型P700) となります。P700+は、PSIIから来る電子により還元され、基底状態のP700が再生します。光合成が抑制される状況になると、P700が酸化され、P700+を蓄積することによってO2へ電子を与えるP700*の存在率を低下させ、ROS生成を抑制します。
研究の内容
マンチェスター大学のGiles Johnson上級講師、および三宅親弘教授は、P700酸化に伴い、PSIで循環的電子伝達反応 (CEF) が誘導されることを見出していました。しかしこれまで、P700酸化とCEFの関係に言及する報告は皆無でした。
本研究では、CEFの分子メカニズムとその性質を明らかにするために、PSI複合体での電子伝達反応にかかわる電子伝達体 (P700、プラストシアニン(PC)、フェレドキシン(Fd))と、リニア電子伝達反応の活性を評価するPSIIの量子収率 [Y(II)] の相互作用を解析しました。実験材料は、主要作物であるコムギ生葉を用いました。ただし、本研究成果の再現性はシロイヌナズナなど多くの植物で検証済みです。
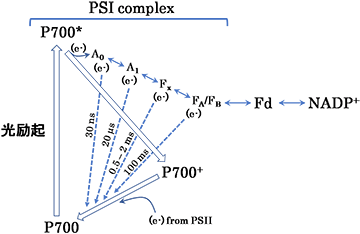
実験の結果、P700*からFdへ電子が流れていく途中には、電子伝達体A0, A1, Fx, FA/FBが存在し、P700+が蓄積する状況では、これら電子伝達体から電子が流れ込む反応 (電荷再結合反応、Charge Recombination) が進行することを明らかにしました (図2)。
P700*は電荷分離し、電子伝達体A0に電子を渡し、自身はP700+へ酸化されます。P700+は、PSIIから来る電子を受け取り基底状態へ還元されます。一方、A0が受け取った電子は、A1, FX, およびFA/FBを渡り、Fdを経てNADP+に流れ、最終的にNADPHが生成されます。P700+が蓄積すると、電子伝達体であるA0, A1, FX, およびFA/FBとP700+との間で電荷再結合反応 (電子の戻り:図2 点線矢印) が生じます。生葉で観測される反応速度時間域からすると、FxとP700+間での電荷再結合反応が有力な経路と考えられます。
電荷再結合反応そのものは、90年代初めより、生化学的に単離されたPSI複合体あるいはラン藻、緑藻などの細胞レベルで、その存在が明らかにされていました。しかし、光合成におけるその生理的役割や、メカニズムなどは未解明のままでした。本研究では、P700+の反応性をもとに、FXからP700+へ電子が流れていることを示唆することが出来ました。
さらに、二つ目の成果として、電荷再結合反応によるROS生成抑制メカニズムを解明しました。電子伝達体A0, A1, FX, FA/FBは、O2一電子還元反応の酸化還元電位と比較して非常に低い酸化還元電位を持ちます。このことは、容易にO2へ電子を与え、ROS生成をもたらすことを示唆します。今回、我々が明らかにした電荷再結合反応は、これら電子伝達体とO2の相互作用を抑制する生理的役割を持っています。
今後の展開
本研究により、P700酸化で誘導されるCEFは、PSI複合体内部で生じる電荷再結合反応であることが提唱されました。
従来の研究では、生葉で観測されるCEFは、Fdに依存したCEFであることが提唱されてきました。しかし、Fdに依存したCEFの反応速度は非常に小さく、さらに、Fd依存CEFにかかわる遺伝子が欠失したシロイヌナズナ変異株では、P700が酸化されると、大きな活性をもつCEFが誘導されます。これまで、この矛盾に対する分子メカニズムは明らかにされていませんでした。
本研究成果は、これらの生理現象に対して、誘導条件および生理状況でのCEFの電子伝達速度を全てうまく説明できるものとなりました。
今後は、PSI複合体内部での電荷再結合反応の生理・生化学、さらに分子生物学的解析によりその役割の普遍性を検討していく必要があります。
用語解説
※1 チラコイド膜
葉緑体がもつ小胞を構成する二重膜。膜内に種々のタンパク質が存在し、光合成の明反応を起こす。
※2 フェレドキシン (Fd)
非ヘム鉄と酸不安定性硫黄を含む、比較的小さな鉄たんぱく質。光合成においては電子伝達系で電子運搬作用を担っている。
※3 クロロフィル
植物細胞に含まれる緑色の色素。光合成の明反応で光エネルギーを吸収する役割をもつ。葉緑素。
謝辞
本研究は、科学技術振興機構 (JST) 戦略的創造研究推進事業 (CREST) 研究領域「環境変動に対する植物の頑健性の解明と応用に向けた基盤技術の創出」(研究総括:田畑哲之) における研究開発課題「活性酸素生成抑制システムの非破壊評価系の確立とフィールドへの応用~危機早期診断システムの構築~」(研究代表者:三宅親弘) の支援を受けて実施されました。
論文情報
タイトル
“Oxidation of P700 Induces Alternative Electron Flow in Photosystem I in Wheat Leaves”
DOI
10.3390/plants8060152
著者
Kanae Kadota1, Riu Furutani1, Amane Makino2, Yuji Suzuki3, Shinya Wada1 and Chikahiro Miyake1*
1 神戸大学農学部
2 東北大学大学院農学研究科
3 岩手大学農学部
* 責任著者掲載誌