神戸大学大学院農学研究科の古谷吏侑 大学院生、三宅親弘 教授らの研究グループは、これまでの研究において明らかにしてきた光合成モデルが植物の内在的な生理現象に対する応答として当てはまることを示すとともに、Rubiscoにより触媒される光呼吸が野外環境において重要な役割を持つことを強く示唆しました。
光合成のモデルに関しては、いまだ多くの議論が行われており、その詳細には不明な部分が多く残っています。本研究グループは近年、C3植物における新しい光合成モデルを見出しており、本研究において、このモデルを実際の光合成に当てはめることができるかを検証しました。
この研究成果は、2020年12月12日付け (現地時間) で、国際科学誌『plants』に掲載されました。
ポイント
- 光合成には、酸化障害をもたらす活性酸素種 (Reactive Oxygen Species, ROS) 生成の危険性が常に伴う。
- これまでの研究から、ROS生成抑制のための “P700酸化モデル” という制御メカニズムが明らかになってきていた。
- イネでは、自身の内在的な生理現象によりROS生成の危険性が高くなりうることが分かったが、“P700酸化モデル” が機能し、安全に光合成反応を進行させていることを明らかにした。
- これまでその生理的意義を疑問視されていた光呼吸が、野外環境での光合成においてP700酸化を通じた重要な機能を果たしていることを強く示唆した。
研究の背景とこれまでの経緯
光合成反応は大きく二つの反応からなります。一つは、光のエネルギーを生体内の反応に利用するために化学エネルギーであるNADPHとATPに変換する光合成電子伝達反応で、もう一つは、化学エネルギーNADPHとATPを用いて、二酸化炭素 (CO2) を有機化合物に固定し、糖を生合成するCO2固定反応です。これら二つの反応系は、エネルギーの供給と消費の関係にあり、そのバランスは保たれる必要があります。
光合成電子伝達系は、葉緑体チラコイド膜※1 上で進行し、膜に存在する光化学系I(PSI)と光化学系II(PSII)が、光エネルギーを吸収します。PSIIでは、光エネルギーを利用し、水を酸化することにより電子、すなわち還元力という化学エネルギーに光エネルギ−が変換されます。このとき、水は酸素と水素イオン(H+)に分解され、酸素は気孔を通じて大気へ、H+はチラコイド膜の内側に放出されます。そして、PSIIで引き抜かれた還元力は、チラコイド膜内にH+を取り込みながらPSIまで伝達されます。PSIでは、PSIIから伝達されてきた還元力を光により再度上昇させ、フェレドキシン(Fd)を介してNADP+を還元し、NADPHを合成します。また、一連の反応によりチラコイド膜の内側に蓄積したH+の濃度勾配(ΔpH:チラコイド膜内外の濃度差)を化学エネルギーとしてATP (アデノシン三リン酸) 合成酵素によるATPが生合成反応を駆動します。
一方CO2固定反応は、葉緑体ストロマ※2 で進行し、酵素Rubisco (Ribulose 1,5-bisphosphate carboxylase/oxygenase) により、葉の表面に存在する気孔から取り込まれ、葉緑体に拡散したCO2がリブロース1,5-二リン酸 (RuBP) に固定され、3-ホスホグリセリン酸 (PGA) を生成します。このPGAから糖生成のための炭素を引き抜きながら、RuBPを再生産するカルビン回路の反応において、電子伝達系からのNADPHおよびATPが消費されます。
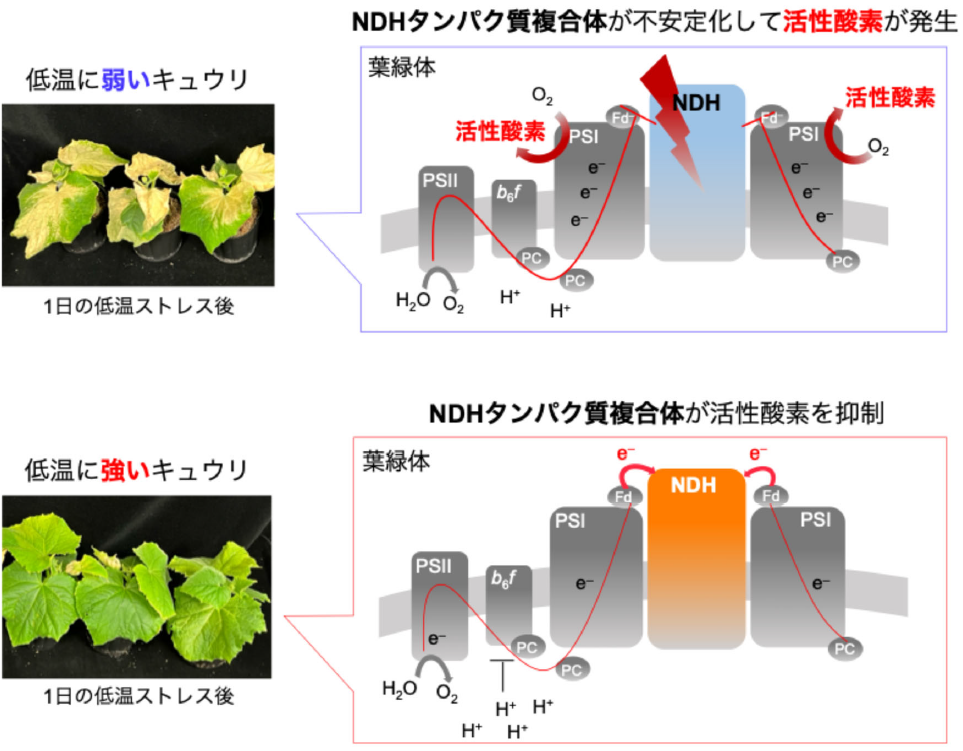
しかし、強光や乾燥などのストレス環境では、CO2固定反応の効率が低下します。先述したように、光合成電子伝達反応とCO2固定反応は密に相互関係する形で機能しています(Furutani et al. 2020, Miyake 2020)。これは、貴重な光エネルギーを光合成に利用するために光合成生物が進化の過程で築きあげてきたものです (Hanawa et al. 2017)。ただし、このことは光合成生物にROSの生成と、生成したROSによる酸化障害の危険性を不可避的にもたらしました。CO2固定反応が滞ると、光合成電子伝達反応の進行が抑制され、光合成電子伝達系に電子が蓄積する可能性が高まります。これまでの研究で、人為的に光合成電子伝達系へ電子を蓄積させると、PSIにおいて酸素が一電子還元され、ROSであるO2 –が生成することが分かっています。このROSは、蓄積するとPSIに酸化障害を与え、光合成能力を低下させます(Sejima et al. 2014)。
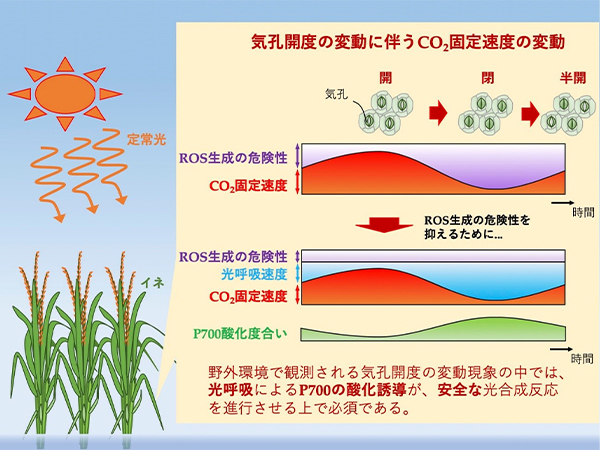
しかし近年、このROS生成を抑制する分子メカニズムが存在することが明らかとなってきました (Sejima et al. 2014, Furutani et al. 2020, Miyake 2020)。これらの研究では、PSIの反応中心クロロフィルであるP700が酸化状態で保たれることにより、ROS発生の場であるPSIでの電子の蓄積を緩和することが分かっています。P700の酸化は、チラコイド膜ルーメンの酸性化 (ΔpH形成の促進) やPQの過還元化による電子伝達の抑制 (RISE; Reduction-Induced Suppression of Electron flow) により生じます。本研究チームは、それまで光合成にとって、光合成を抑制し生長性を低下させる悪者と言われてきた“光呼吸代謝”がP700酸化、ひいては酸化障害を防ぐためになくてはならないことを明らかにし、新しい光合成のモデルの確証を得るに至りました。
研究の内容
しかしこれまでの研究は、環境制御された人為的ストレスに対する光合成応答を解析したものであり、野外環境における「P700酸化モデル」の検証は未着手でした。そんな中、イネでは一定の光環境下においても、30~60分の周期で最大5倍近くCO2固定反応速度が変動する現象を観測しました (Furutani et al. 2020)。この変動は、気孔の不安定な開閉によってもたらされていることも分かりました。気孔開度の変動現象は、野外環境において、さまざまな植物種で報告がなされています。そこで、この連続的かつ内在的な光合成変動の生理現象を、光合成における「P700酸化モデル」により説明できると考え、これを検証する実験を行いました。
本研究では、気孔開閉の周期的変動によるCO2固定反応速度の変動に対して、光呼吸反応がCO2固定反応の低下を埋め合わせるようにその活性が上昇することで、光合成電子伝達系の電子を安定して消費し、PSIから電子を受容するFdの酸化レベルをほぼ一定に保つことを明らかにしました。この光呼吸は、安定した電子の消費を行うだけではなく、ATP合成反応によるチラコイド膜ルーメンのプロトンの消費を抑えることでΔpHを誘導しており、ΔpH形成促進によるP700酸化に大きく寄与していました。この光呼吸を抑制する低酸素条件 (2 kPa O2) では、CO2固定速度の変動に合わせてPSIIでの電子伝達速度、ΔpHが大きく変動しました。しかしながら、P700の酸化度は光呼吸駆動時と比べて、小さくなっていました。一方、P700酸化の抑制が緩和されるフェーズにおいては、PQの還元状態が高く維持されることを見出しました。このことから、光呼吸とCO2固定反応の両方が抑制されるような状況では、RISEがP700酸化に寄与しうることを明らかにしました。
本研究は、これまで見出されてきたP700酸化を中心とした光合成モデルが、植物体自身の内在的な生理現象の中でも成立することを示すとともに、野外環境の光合成における普遍的な光呼吸の重要性をさらに強調する成果となりました。
今後の展開
現在、光呼吸がなぜプロトンの消費速度を抑制し、ΔpHを誘導することができるかについての詳細なメカニズムは明らかではありません。今後、光呼吸とΔpHの関連に関して詳細な解析が求められます。また、P700酸化をもたらすための電子伝達制御メカニズムについても、未知のものが存在すると考えられ、その分子的実体の解明を行う予定です。
今回の研究は光合成生物の中でも、被子植物、またその中のC3植物での光合成のモデルの検証になります。今後、シアノバクテリアやコケ植物、シダ植物、裸子植物、C4植物といった様々な植物種での光合成モデルの解明と、進化との関連付けを目指し、解析を進め、P700酸化メカニズムの多様性、そして光呼吸の存在意義の解明を行っていく予定です。
用語解説
- ※1 チラコイド膜
- 葉緑体がもつ小胞を構成する二重膜。膜内に種々のタンパク質が存在し、光合成の明反応を起こす。
- ※2 ストロマ
- 葉緑体の主要な区画。グラナ (円板の形をした緑色の膜構造体) の周囲にあり、無色の液体状をしている。Rubiscoが構成タンパク質の約半分を占める。
謝辞
本研究は、以下の支援を受けて行われました。
科学技術振興機構 (JST) 戦略的創造研究推進事業 (CREST)
研究領域「環境変動に対する植物の頑健性の解明と応用に向けた基盤技術の創出」(研究総括:田畑 哲之)
研究課題名「活性酸素生成抑制システムの非破壊評価系の確立とフィールドへの応用」(JPMJCR15O3) (研究代表者:三宅親弘)
論文情報
- タイトル
- “Intrinsic Fluctuations in Transpiration Induce Photorespiration to Oxidize P700 in Photosystem I”
- DOI
- 10.3390/plants9121761
- 著者
- 古谷吏侑1、 牧野周2、 鈴木雄二3、 和田慎也1、 嶋川銀河4、 三宅親弘1
1 神戸大学大学院農学研究科
2 東北大学大学院農学研究科
3 岩手大学農学部
4 大阪大学大学院工学研究科 - 掲載誌
- 『plants』Volume 9, Issue 12 (December 2020)