東京大学大学院理学系研究科附属臨海実験所の三浦徹教授、小口晃平准教授、千代田創真大学院生、神戸大学大学院理学研究科生物学専攻の青沼仁志教授による研究グループは、都市公園などで容易に採集されるマクラギヤスデを用いて、脱皮(注1)時に体節(注2)が追加される過程を詳細に明らかにしました。
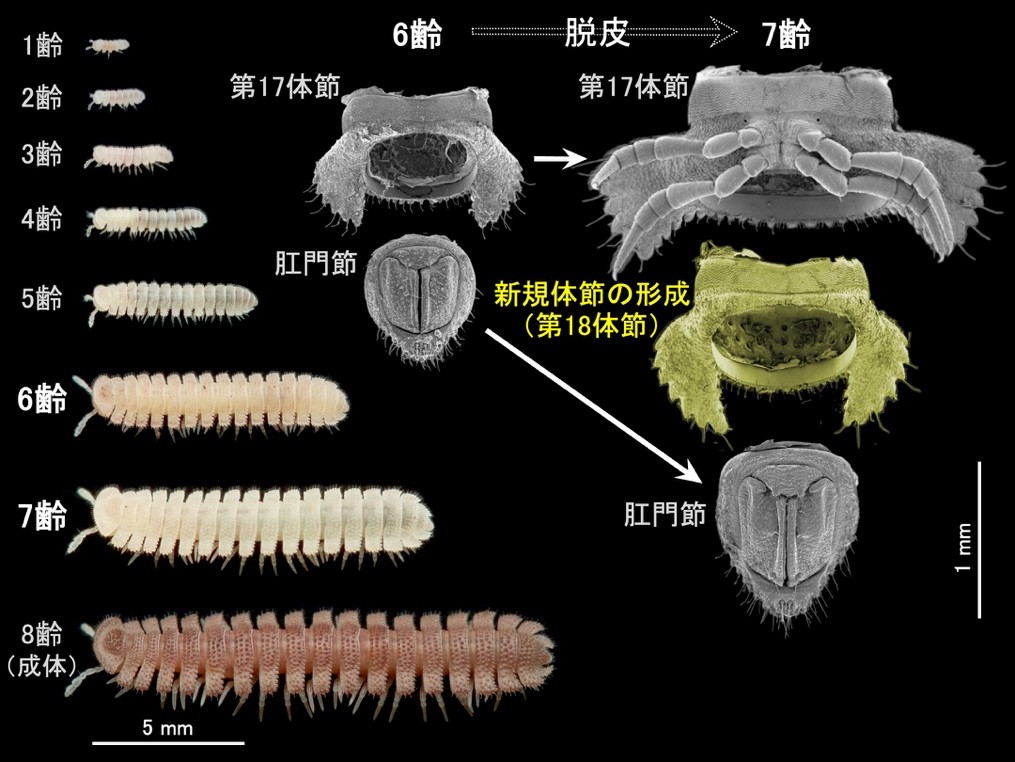
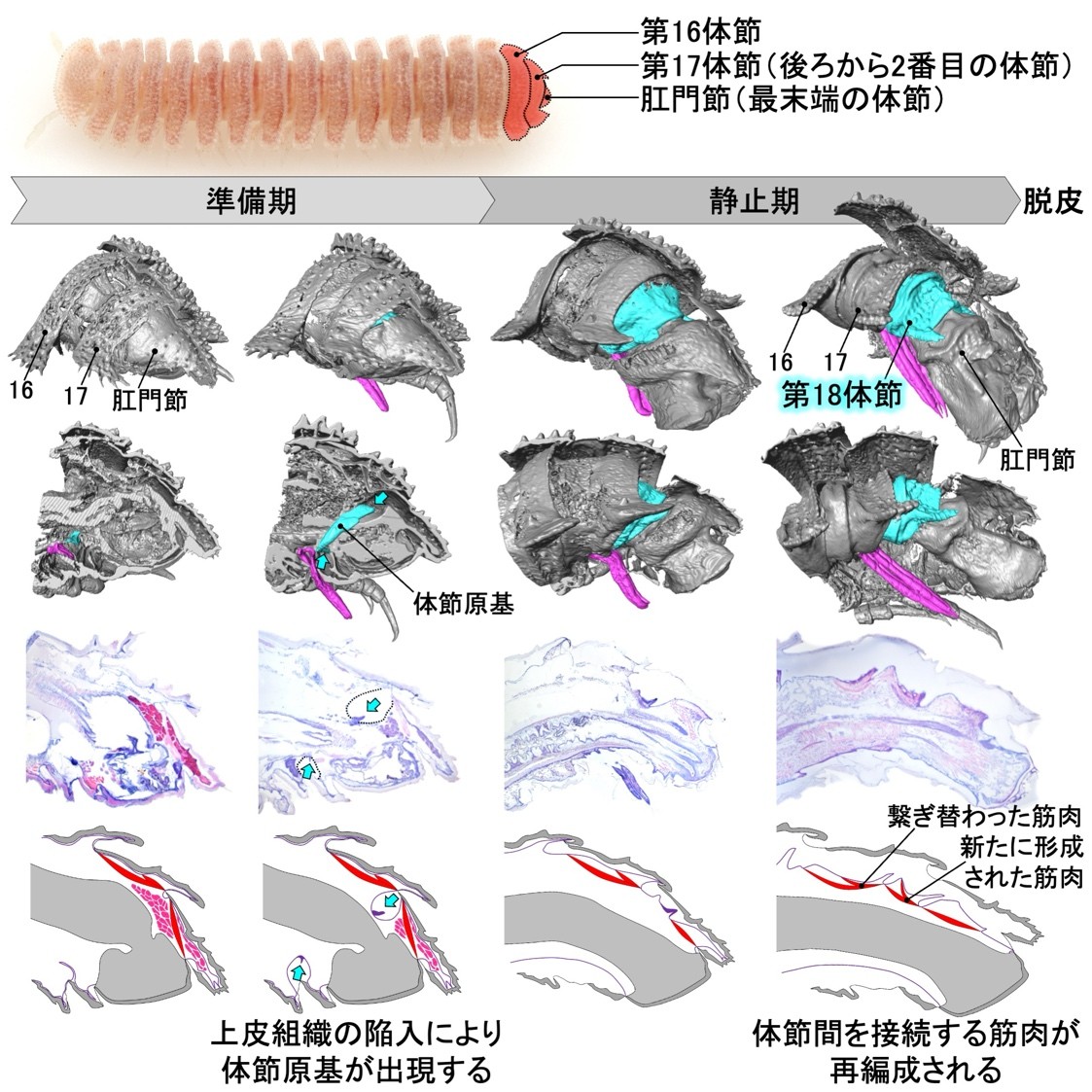
節足動物門(注3)多足亜門(注4)ヤスデ綱(以下、ヤスデ)では、脱皮のたびに体節を増やしながら成長する「増節変態(注5)」という発生様式が見られます(図1)。ヤスデの増節変態では、新たな体節は最末端の肛門節と後ろから2番目の体節の間に追加されることが知られていましたが、どのような形態形成が生じることによって新規体節が形成・追加されるのかは長らく未解明でした。本研究は、行動観察に基づいて脱皮前の状態を定義し、X線マイクロCT(注6)などを用いて各段階における内部構造を詳細に観察することで、肛門節と後ろから2番目の体節の間の上皮組織(注7)が脱皮前に急激に陥入してくびれることで「体節原基」が生じることを明らかにしました。さらに、こうした新規体節の挿入に伴って、前後の体節を接続する筋肉が繋ぎ替わることが示唆されました。
本研究により、増節変態における体節の追加機構を研究する上での基盤となる、体節の追加に伴う組織の変化過程が明らかになりました。今後、こうした現象の背景にある生理発生機構を明らかにすることにより、これまでほとんど胚発生に限られていた節足動物の体節形成機構に関する理解を個体発生全体に拡張するとともに、祖先的と考えられている増節変態から、どのようにして増節変態をしない昆虫類などが現れたのかといった進化過程についても理解が深まることが期待されます。
ポイント
- 脱皮のたびに体節を増やす「増節変態」を行うヤスデにおいて、脱皮前に上皮組織が急激に陥入してくびれることで「体節原基」が生じること、同時に筋肉の再編成が起こることを示した。
- 行動観察に基づいて脱皮前の状態を定義し、X線マイクロCTなどを用いた3次元的な観察を行うことで、新規体節が形成される形態形成過程を初めて詳細に明らかにした。
- 本研究の結果は、増節変態の生理発生機構の一端を明らかにするものであり、節足動物における体節形成の共通性・多様性や進化過程の理解が深まることが期待される。
研究の背景
節足動物は前後軸に沿って並んだ「体節」を持ち、その形成過程は主に昆虫であるショウジョウバエの胚発生を用いた研究から詳細に解明されてきました。昆虫以外の節足動物では、胚発生のみならず孵化後の後胚発生過程においても体節形成を継続する「増節変態」という発生様式も知られています。増節変態は、節足動物における祖先的な発生様式と考えられており、節足動物の生涯を通じた体節形成機構を解明し、その節足動物全体での進化を議論する上で重要な現象です。
ムカデやヤスデなどを含む多足類は、多数の同型の体節が連なった体制が特徴的なグループです。なかでもヤスデは全ての種が増節変態によって成長し、体節追加パターンとして、「新たな体節は脱皮時に最末端の肛門節と後ろから2番目の体節の間に追加される」ことがすでに知られていました。しかしながら、どのような組織の変化によって新たな体節が追加されるのかといった形態形成過程は、これまでほとんど未解明でした。
研究の内容
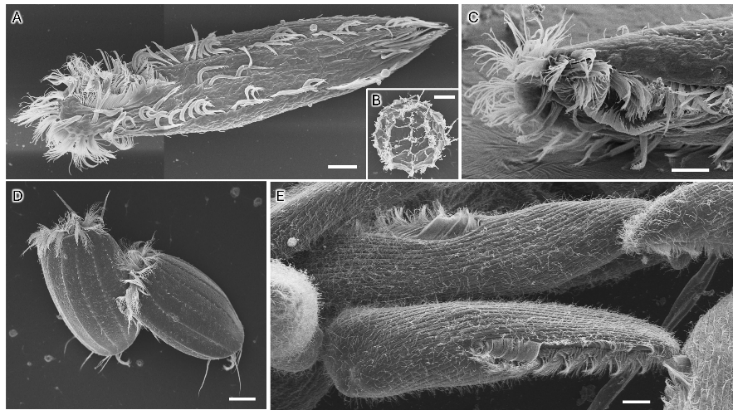
本研究は、ヤスデ綱オビヤスデ目シロハダヤスデ科に属するマクラギヤスデNiponia nodulosaを対象に、脱皮に先立つ内部構造の変化を詳細に観察することで、体節追加に伴う組織レベルの形態形成過程の解明を試みました。
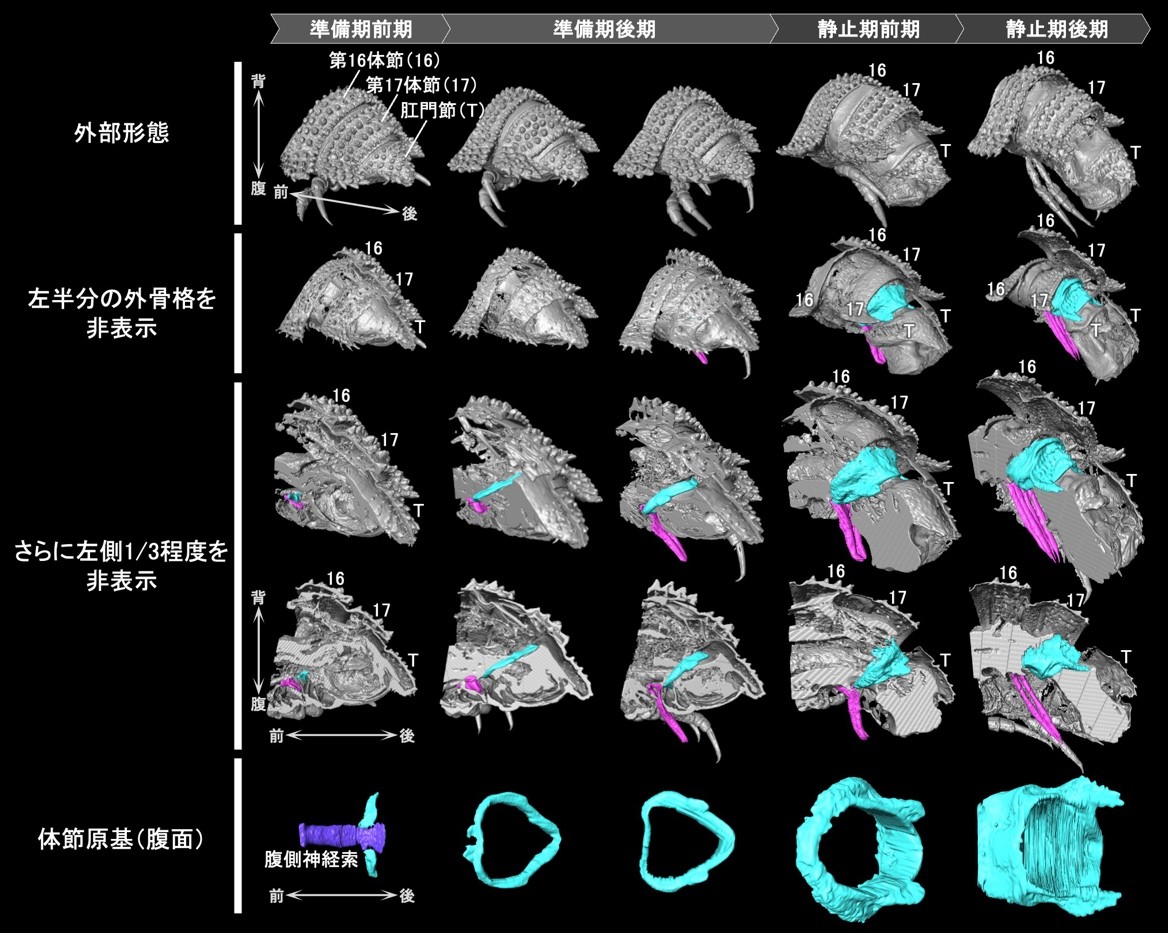
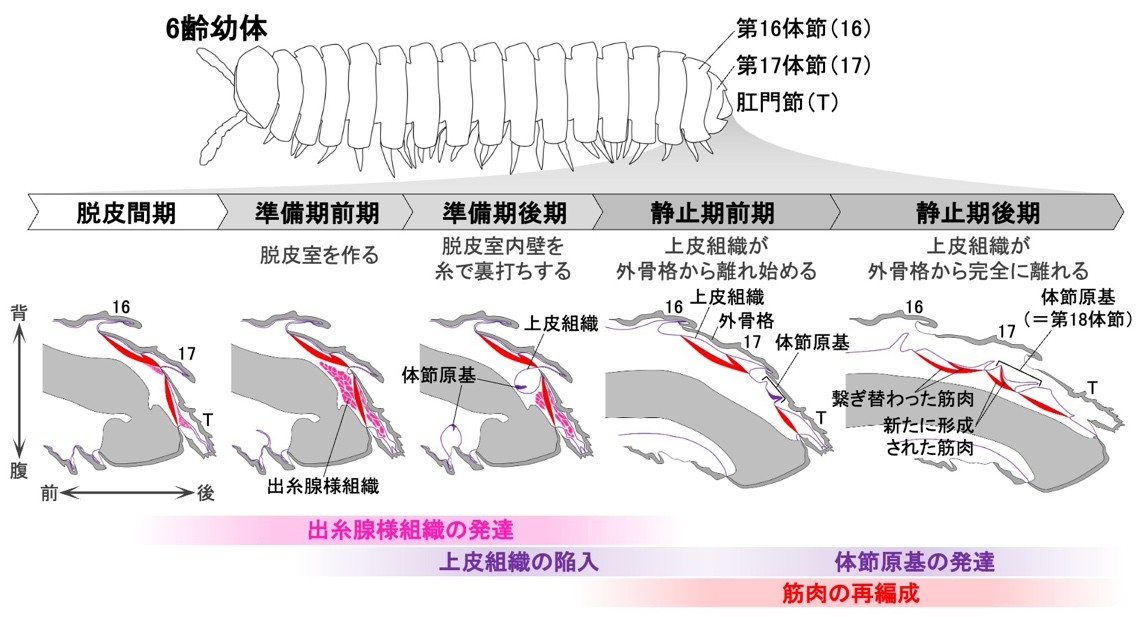
まず、形態形成過程の観察に先立って、脱皮前の詳細な行動観察を行いました。本種の幼体は、脱皮前の「準備期」に「脱皮室」と呼ばれる部屋をつくり、その中で脱皮をします。こうした行動を詳細に観察し、「準備期前期」と「準備期後期」を新たに定義しました(図2)。また、続く「静止期」についても、外部から観察可能な形態変化に基づいて「静止期前期」と「静止期後期」を定義しました。これによって、脱皮前の個体の状態を正確に把握することが可能になりました。

次に、新たに定義されたステージに基づいて、体節の追加が起こる胴部末端の内部形態の変化過程を明らかにするためにX線マイクロCTを用いた3次元的な観察を行いました。その結果、準備期後期にリング状の体節原基が形成され、これが静止期の間に前後方向に大きく発達することが明らかになりました(図3)。
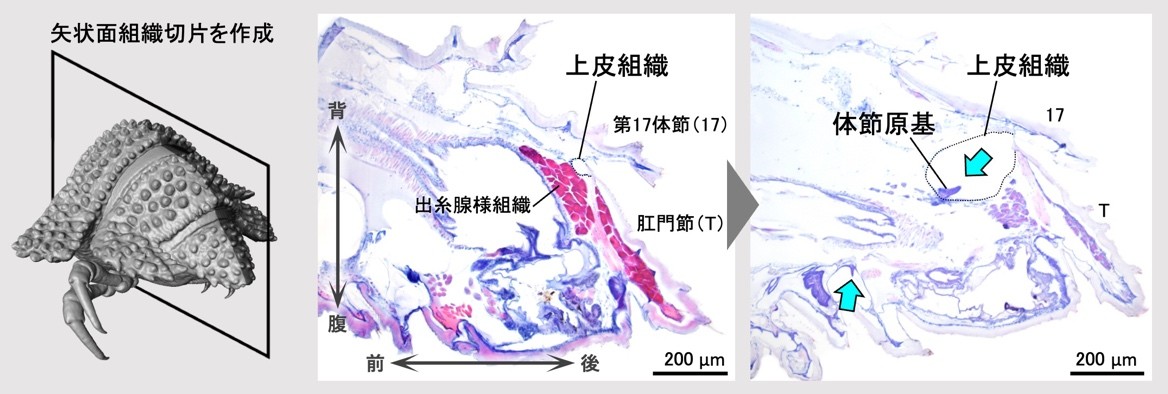
さらに、体節原基の形成・発達の際に起きている組織の変化を明らかにするため、組織学的な観察を行いました。上皮組織に着目した組織切片の観察の結果、準備期後期に、肛門節と後ろから2番目の体節の間で上皮組織が急激に陥入し、陥入部分に体節原基が形成されることがわかりました(図4)。なお、準備期後期には肛門節内部にエオシン(注8)で赤く濃染される組織が発達していましたが、組織の性状や、準備期後期に肛門節末端から糸を出して脱皮室を裏打ちすることから、これは出糸腺(注9)である可能性があります。
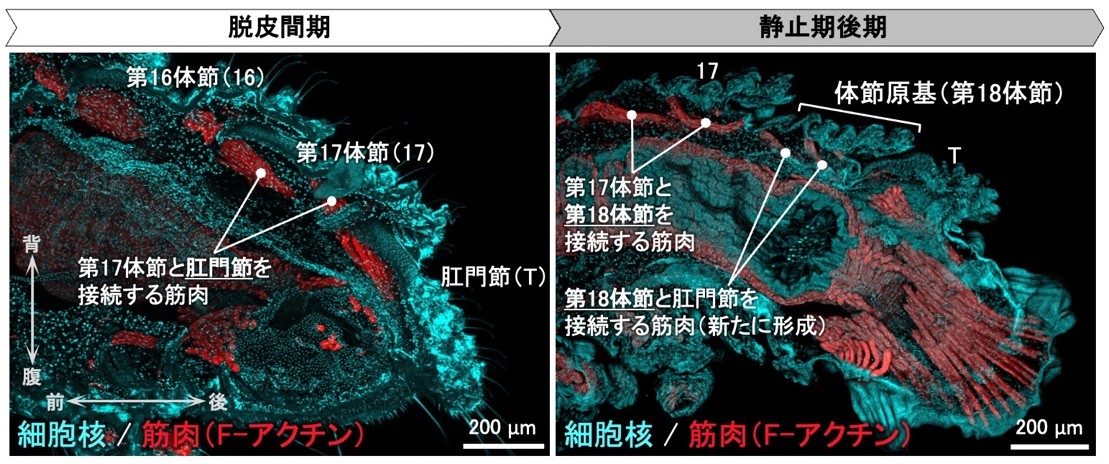
体節の追加に伴う筋肉の挙動についても明らかにするため、共焦点レーザー顕微鏡(注10)を用いた筋肉の蛍光観察を併せて行いました。その結果、後ろから2番目の体節と最末端の肛門節を接続する背側の筋肉は、体節の追加に伴って、肛門節から新たに追加される体節(体節原基)へと繋ぎ替わることがわかりました(図3、図4)。さらに、新たに追加される体節の内部には、その体節と肛門節を接続する筋肉が脱皮に先立って新たに形成されることが示唆されました(図5)。
以上の結果から、マクラギヤスデでは体節の追加に伴って、上皮組織の陥入による体節原基の形成と、筋肉の繋ぎ替えと新生による再編成が脱皮前の胴部末端で一過的かつ局所的に生じることがわかりました(図6)。
研究の展望
本研究により、増節変態において、「いつ」「どこで」「何が」起こることで体節の追加が起こるのかといった基本的な形態形成過程が明らかになりました。私たちの研究グループによるこれまでの研究(関連情報参照)と併せて、未だほとんど明らかになっていない増節変態のメカニズムに迫る上で基盤となる、詳細な組織形態学的知見が整理されました。
マクラギヤスデは、東京大学本郷キャンパスにも生息しているとても身近な生き物ですが、地道な形態・行動の観察とX線マイクロCTなどの最新技術を組み合わせることで、その知られざる一面が明らかになりました。今後は、こうした現象の背景にある生理発生機構をさらに詳細に明らかにしていくことで、胚発生における体節形成と増節変態の共通性、あるいはそれぞれの特異性について理解が深まることが期待されます。さらに、節足動物において祖先的と考えられている増節変態から、どのようにして増節変態をしない昆虫などが現れたのかといった進化過程について議論することで、節足動物の多様化のメカニズムの一端に迫れると期待されます。
用語解説
(注1)脱皮
昆虫や甲殻類などの節足動物において、成長する過程でクチクラからなる硬い外骨格を脱ぎ捨てる現象のこと。
(注2)体節
節足動物の体を構成する基本単位で、通常1体節は1対の脚を備える。しかし、ヤスデは「倍脚類」とも呼ばれるように例外的で、見かけ上の1体節が2対の脚を備えている。このことから、ヤスデの見かけ上の1体節は発生学的には2体節分とみなすべきであるが、ここでは単に見かけ上の体節を1体節として扱っている。
(注3)節足動物門
昆虫、甲殻類、多足類、鋏角類を含む動物門で、最も多様性の高い動物門である。昆虫は増節変態をせず、全ての種が整形変態である。一方他の3つのグループでは、増節変態と整形変態のどちらも見られる(注5参照)。
(注4)多足亜門
節足動物門に属する亜門の1つで、「多足類」と呼ぶこともある。現生種は全て陸生である。多くの種は細長い体型をしており、多数の同規的な体節と脚を持つ。ムカデ、ゲジ、ヤスデなどを含む。
(注5)増節変態
節足動物において、脱皮のたびに体節を増やしながら成長する発生様式のこと。反対に、脱皮ごとに体節を増やさずに成長する発生様式を「整形変態」という。現生の節足動物の系統関係や化石記録などから、増節変態が祖先的であると考えられている。
(注6)X線マイクロCT
X線を用い、試料の内部構造を1 µm程度の高い分解能で非破壊で3次元的に可視化する技術。
(注7)上皮組織
上皮細胞からなるシート上の組織で、節足動物の場合はクチクラからなる外骨格を裏打ちして、その形成や維持を担う。
(注8)エオシン
組織切片の染色に用いられる色素で、タンパク質が豊富に含まれる細胞質などを赤色に染める。
(注9)出糸腺
糸の材料となるタンパク質などを合成・貯蔵する組織で、そこから体外に分泌された物質が糸として機能する。節足動物ではクモやカイコなどでよく知られ、「糸腺」「絹糸腺」と呼ばれることもある。
(注10)共焦点レーザー顕微鏡
試料にレーザー光を照射して、試料から発する蛍光を検出することによって画像を取得する顕微鏡の一種。タンパク質などの生体分子を蛍光標識したサンプルを観察することで、特定の構造を可視化することができる。
研究助成
本研究は、科研費「特別研究員奨励費(課題番号:24KJ0891、千代田創真)」、「基盤研究(A)(課題番号:25H00988、三浦徹)」の支援により実施されました。
論文情報
タイトル
DOI
10.1186/s13227-025-00257-3
著者名
Soma Chiyoda, Kohei Oguchi, Hitoshi Aonuma, Toru Miura* (*責任著者)
掲載誌
Developmental Biology Advances