神戸大学大学院農学研究科の高田紗都子 (博士前期課程修了) とアクタ アヤシャ (博士後期課程) らは、ハクサイ、コマツナ、カブ等のアブラナ科葉根菜において、低温処理前の開花抑制遺伝子FLCの発現量が、これらの葉根菜の開花を抑制するのに重要であることを示しました。今後、アブラナ科葉根菜の品種育成への貢献が期待されます。
本研究は、神戸大学大学院農学研究科の藤本龍准教授ら、新潟大学大学院自然科学研究科岡崎桂一教授、シェイ ダニエル (研究員)、農研機構、オーストラリア連邦科学産業研究機構 (CSIRO) らの研究グループにより行われました。
この研究成果は、2019年9月25日午前10時 (イギリス時間) に、Scientific Reportsにオンライン掲載されました。
ポイント
- アブラナ科葉根菜の開花に必要な遺伝子の機能を明らかにした。
- アブラナ科葉根菜の春化※1の鍵遺伝子として知られているFLOWERING LOCUS C (FLC) の低温処理前の発現量が、低温による花成誘導 (花を咲かせる) の抑制に重要であることを明らかにした。
- ハクサイ、コマツナ、カブ等のアブラナ科葉根菜は、花芽をつけると商品価値が損なわれることから、栽培時に低温にさらされても花成誘導されにくい品種 (晩抽性品種) への需要が高く、本研究成果によるアブラナ科葉根菜の品種育成への貢献が期待される。
研究の背景

アブラナ科野菜であるハクサイ、コマツナ、カブ等のBrassica rapa種 (図1) は、花成誘導のためには、一度長期間の低温にさらされる必要があります。このような現象を春化と呼びます。B. rapa種の中には、比較的短い期間の低温にさらされただけで花を咲かせる系統や、非常に長い期間低温にさらされないと花を咲かせることができない系統が存在します。この両者の違いがどのような分子機構で説明できるのかは完全には明らかにされていません。
アブラナ科葉根菜は栽培中に花芽が形成されると、薹立ち (とうだち)※2によって商品価値が損なわれます。そのため、低温にさらされても花芽が形成されにくい (薹立ちしにくい) 晩抽性品種が育成されています。つまり、アブラナ科葉根菜の品種育成においては、春化メカニズムの理解が重要となります。
研究の内容
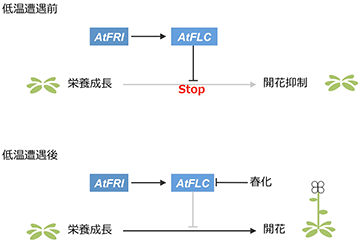
灰色の線はその経路が機能していないことを示す。
これまでの研究で、モデル植物のシロイヌナズナ (アブラナ科)では、低温にさらされる前は、FRIGIDA (AtFRI) 遺伝子により開花抑制遺伝子AtFLCの発現が誘導されることで、花成が抑制されており (図2上)、低温にさらされると、AtFLC遺伝子の発現が抑制され、花成が誘導される (図2下) ことが分かっています。
本研究の研究対象であるB. rapa種には、2つのFRI遺伝子(BrFRIa、BrFRIb) と4つのFLC遺伝子 (BrFLC1、BrFLC2、BrFLC3、BrFLC5) が存在します。まず、B. rapa種のこれらの遺伝子の機能を調べました。
AtFRIタンパク質が機能していないため、AtFLC遺伝子の発現が誘導されず早咲きとなるシロイヌナズナに、B. rapa種のBrFRIbを遺伝子導入したところ、得られた形質転換体は遅咲きになりました。また、この形質転換体ではシロイヌナズナのAtFLC遺伝子の発現が誘導されていました。このことから、BrFRIbタンパク質には、FLC遺伝子の発現を促進する機能があることが示されました (図3)。
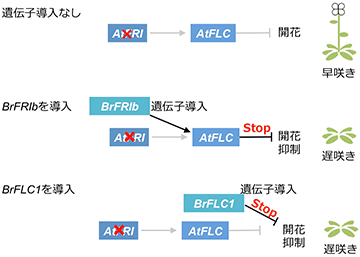
灰色の線はその経路が機能していないことを示す。×印は遺伝子が機能していないことを示す。BrFLC2とBrFLC3を使用した実験では、BrFLC1の結果と同様の結果が得られた。
前述の実験に使用したものと同じシロイヌナズナの系統に、B. rapa種のBrFLC1、BrFLC2、BrFLC3をそれぞれ遺伝子導入した形質転換体を作出したところ、全て遅咲きとなる形質転換体が得られました。このことから、BrFLC1、BrFLC2、BrFLC3は全て開花抑制遺伝子であることが明らかとなりました。BrFLC5は偽遺伝子※3であるため、今回の解析からは除いています。今までのB. rapa種の春化に関する研究では、個別のBrFLC遺伝子の機能と開花との関連性を示唆するものが主流でした。しかし、我々は、春化においては、3つの機能的なBrFLC遺伝子全てを考慮する必要性があると考えました。
B. rapa種の10系統について、低温処理前のBrFLC1、BrFLC2、BrFLC3、BrFLC5遺伝子の発現量の総量、および、低温処理 (4℃) を4週間行った後に、22℃で生育させた時、花芽をつけるまでに要する日数を調べました。すると、BrFLC遺伝子の発現量の総量と開花日数には、正の相関関係が見られました (図4)。このことから、低温処理前のBrFLC遺伝子の発現量の総量が低温による花成誘導の起こりにくさにとって重要である可能性が明らかになりました。しかし、今回調べた10系統のうち1系統 (BRA2209) は、低温処理前のBrFLC遺伝子の発現量の総量は多くないにもかかわらず、低温処理後の花芽誘導にかかる日数は長くなりました (図4)。この系統を調べたところ、低温処理を行っても、BrFLC遺伝子の発現抑制が起こりにくいことが分かりました (図5)。よって、低温処理によるBrFLC遺伝子の発現抑制が起こりにくいという特徴も、低温による花芽誘導の起こりにくさという点において重要となることが明らかとなりました。
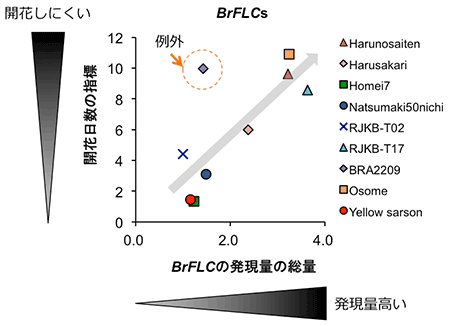
BRA2209系統は、BrFLC遺伝子の発現量の総量が低いが遅咲きとなり例外的。
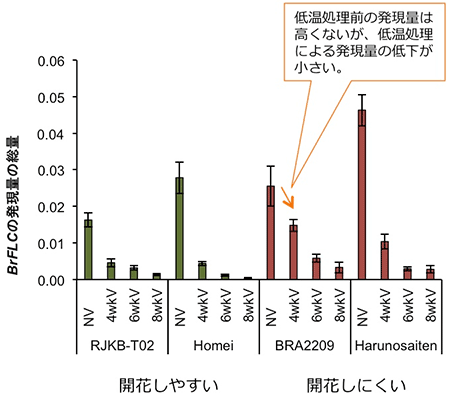
4、6、8wkVは、それぞれ4、6、8週間の低温処理を示す。
本研究から、ハクサイ、コマツナ、カブ等のB. rapa種の低温による花成誘導を防ぐためには、低温処理前のBrFLC遺伝子の発現量の総量を高くし、さらに、低温処理によっても、BrFLC遺伝子の発現が抑制されない対立遺伝子を利用することが重要であることが示されました。
今後の展開
今回の研究から、ハクサイ、コマツナ、カブ等のB. rapa種では、低温処理前に高い発現量を示し、かつ、低温処理によって発現抑制が起こりにくいBrFLC遺伝子を利用することで、低温に遭遇しても花成誘導が起こりにくい晩抽性品種が育成できると考えられます。今後はこの可能性を実証するために、理想的なBrFLC遺伝子を持たせた場合に、晩抽性を示すかを検証することで、これらアブラナ科葉根菜の晩抽性品種育成に貢献できればと考えています。
用語解説
※1 春化
長期間の低温に遭遇することによって花成が誘導されること。
※2 薹立ち
花成誘導により、花茎が伸びだすこと。抽だいともいう。葉菜類の多くは、葉が固くなって食味が落ちてしまう。
※3 偽遺伝子
かつては遺伝子として機能していたが、変異により現在は機能を失っている遺伝子
謝辞
本研究は、以下の支援を受けて行われました。
- 日本学術振興会 二国間共同研究事業
- 日本学術振興会 国際共同研究加速基金
- 日本学術振興会 科学研究費 基盤研究B (15H04433, 18H02173)
- ひょうご科学技術協会 学術研究助成事業
論文情報
タイトル
“The role of FRIGIDA and FLOWERING LOCUS C genes in flowering time of Brassica rapa leafy vegetables”
DOI
10.1038/s41598-019-50122-2
著者
Satoko Takada+, Ayasha Akter+, Etsuko Itabashi, Namiko Nishida, Daniel J. Shea, Naomi Miyaji, Hasan Mehraj, Kenji Osabe, Motoki Shimizu, Takeshi Takasaki-Yasuda, Tomohiro Kakizaki, Keiichi Okazaki, Elizabeth S. Dennis, Ryo Fujimoto
+ These authors equally contributed to this work掲載誌









