理化学研究所 (理研) 生命機能科学研究センター染色体分配研究チームの三品達平基礎科学特別研究員 (研究当時、現客員研究員)、京都大学生態学研究センターの佐藤拓哉准教授、国立台湾大学の邱名鍾助教、大阪医科薬科大学医学部の橋口康之講師 (研究当時)、神戸大学理学研究科の佐倉緑准教授、岡田龍一学術研究員、東京農業大学農学部の佐々木剛教授、福井県立大学海洋生物資源学部の武島弘彦客員研究員らの国際共同研究グループは、ハリガネムシのゲノムにカマキリ由来と考えられる大量の遺伝子を発見し、この大規模な遺伝子水平伝播[1]がハリガネムシによるカマキリの行動改変 (宿主操作[2]) の成立に関与している可能性を示しました。
本研究成果は、寄生生物が系統的に大きく異なる宿主の行動をなぜ操作できるのかという謎を分子レベルで解明することに貢献すると期待されます。
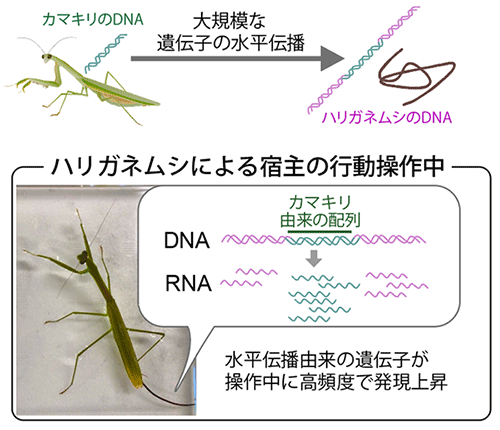
自然界では、寄生生物が自らの利益のために宿主操作を行う例が数多く確認されています。今回、国際共同研究グループは、寄生虫ハリガネムシと、寄生により入水行動をさせられる宿主カマキリの全遺伝子発現 (トランスクリプトーム[3]) 解析を行いました。宿主操作に伴う明瞭なトランスクリプトーム変化はカマキリではなくハリガネムシのみに見られたことから、寄生虫によって生合成された分子が宿主の行動操作に関わっている可能性が示唆されました。さらに、発現量が変化したハリガネムシ遺伝子には、宿主であるカマキリ遺伝子とDNA塩基配列レベルで非常によく似ているものがより多く含まれており、それらの遺伝子の中には、カマキリの行動操作に関係し得る機能を持つものが見いだされました。これらの結果から、ハリガネムシは、宿主であるカマキリから大規模な遺伝子水平伝播を受けることで、宿主操作を成し遂げている可能性が明らかになりました。
本研究は、科学雑誌『Current Biology』オンライン版 (10月19日付:日本時間10月20日) に掲載されました。
背景
今日、地球上に生息する生物種の約40%は寄生生物であり、すべての野生動物は少なくとも1種の寄生生物に寄生されているといわれています。動物の行動は通常、個体の生存や繁殖に有利になるように制御されていると考えられています。しかし、野生動物に見られる多様な行動は、実は寄生生物の影響を強く受けて制御されている可能性があります。この顕著な例として、寄生生物が自らの利益 (感染率や繁殖成功率の向上) のために、宿主である野生動物の行動を改変 (宿主操作) する現象が数多く確認されています。寄生生物による宿主操作は、ある生物の遺伝子が別の生物の表現型として発現する「延長された表現型[4]」の好例として、多くの生物学者を魅了し、近年その分子機構が解明されつつあります。しかし、そもそも寄生生物が系統の大きく異なる宿主の生体システムに介入するためのツール (遺伝子群) をどのように手に入れているのかは、ほとんど明らかになっていません。
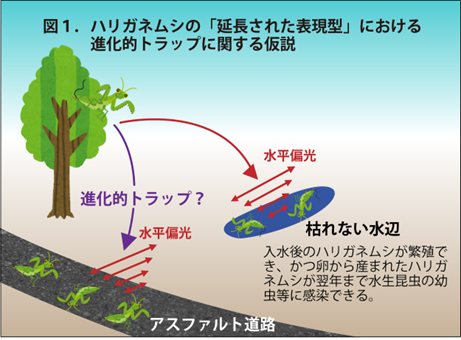
宿主操作の代表例として、陸生昆虫に寄生中のハリガネムシが、交接・産卵場所である川や池に移動するために、寄生相手であるカマキリやカマドウマなど (以下、宿主) を川や池に自ら入水させるという現象が知られています (図1) 注1) 。ハリガネムシに寄生された宿主の入水行動については、100年以上も前から世界各地で報告されていましたが、その分子機構は謎に包まれていました。
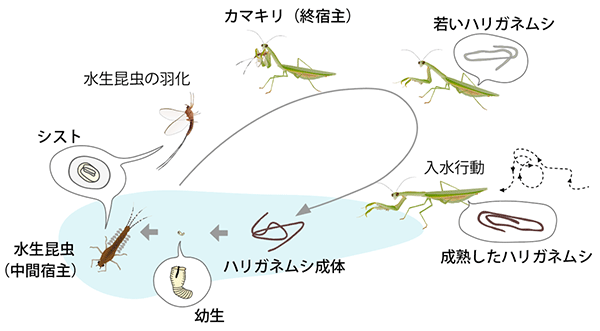
水中で孵化したハリガネムシ幼生は、水生昆虫に寄生してその体内でシスト (休眠状態の形態) になる。水生昆虫の羽化とともに水域から陸域に移動し、水生昆虫がカマキリに捕食されると、その体内で成長する。カマキリの体内で成虫になると、宿主操作して入水させる。カマキリが入水すると、脱出して水域に戻り、繁殖して一生を終える。
注1) 2021年6月22日神戸大学プレスリリース「ハリガネムシは寄生したカマキリを操作し水平偏光に引き寄せて水に飛び込ませる」
研究手法と成果
国際共同研究グループはまず、寄生生物ハリガネムシ (Chordodes fukuii) とその宿主チョウセンカマキリ (Tenodera angustipennis) をモデルとして、宿主操作に際して両者の遺伝子発現がどのように変化しているかを調べました。ハリガネムシについては「操作前」「操作中」「操作後」の個体、カマキリについては「操作前」「操作中」「非感染」の個体のそれぞれ3群に分けて、ハリガネムシの体組織全体とカマキリ全脳のトランスクリプトーム解析を行いました。
その結果、ハリガネムシでは、「操作中」に発現する遺伝子の種類や量は「操作前」や「操作後」と比べて大きく異なっており (図2A)、多数の遺伝子が操作中に特異的な発現変化を示していました (発現上昇:3,168遺伝子、発現低下:1,688遺伝子) 。一方、カマキリの脳内では、操作の前後や感染の有無による遺伝子発現の特徴的な変化は観察されず (図2B)、また、操作中に発現変化する遺伝子もごく少数でした (発現上昇:1遺伝子、発現低下:8遺伝子) 。宿主操作に伴う遺伝子発現変化がハリガネムシのみに顕著に見られたという結果は、ハリガネムシが生合成する分子が宿主であるカマキリの生体システムに介入することで、行動操作を達成していることを示唆します。他の寄生虫やウイルスでは、宿主の生合成する分子と類似する分子 (分子擬態[5]) を生合成することで効果的な介入を実現している例が報告されており注2) 、ハリガネムシにおいても同様の分子擬態を利用している可能性が考えられます。
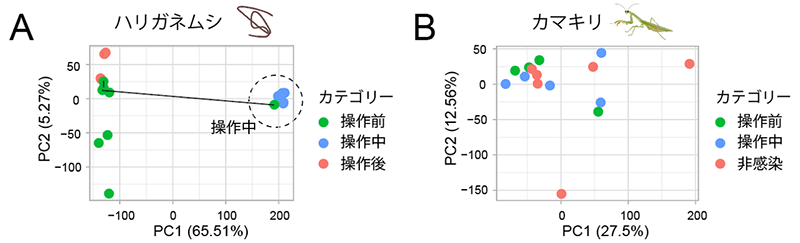
ハリガネムシの全体組織 (A) とカマキリの脳 (B) のトランスクリプトーム解析から得た遺伝子発現プロファイル (遺伝子発現全体の傾向) を、統計的手法 (主成分分析) により二つの変数 (PC1:主成分1、PC2:主成分2) に縮減し、2次元グラフで表現したもの。点はそれぞれ操作前 (緑)、操作中 (青)、操作後 (赤:ハリガネムシ)、非感染 (赤:カマキリ) の個体。本実験では偶然、1匹のカマキリから2個体のハリガネムシが得られた例があり (多重感染)、そのハリガネムシのペアの点を線でつないで表現している。ハリガネムシ (A) では、操作中のトランスクリプトーム変化に主成分1が大きく寄与している (寄与率65.51%) が、カマキリ (B) ではカテゴリー間で明瞭なパターンは見いだせなかった。
そこで、ハリガネムシの合成する分子の中にカマキリの分子擬態をしているものがあるかを検証するため、実験に用いたChordodes属ハリガネムシとは属レベルで異なる複数のハリガネムシ種や、その宿主種を広く含むアミノ酸およびDNA配列から成る生物網羅的な遺伝子データベースを構築し、種間で配列の類似した遺伝子がどの程度存在するかを検索しました。その結果、Chordodes属ハリガネムシの遺伝子の大半は、他属ハリガネムシの遺伝子とアミノ酸配列の高い一致度を示しました。しかし驚くべきことに、系統的に大きく異なる宿主カマキリの遺伝子とも、アミノ酸配列のみならず (図3A)、DNA塩基配列レベルでも高い類似性を有する遺伝子が多数あることが分かりました (図3B) 。つまり、ハリガネムシは、一部の分子の生合成においてカマキリの分子擬態をしている可能性が高いといえます。

(A) ハリガネムシの遺伝子 (アミノ酸配列) に対して最も相同性が高い種を生物網羅的に検索し、属単位で該当するハリガネムシの遺伝子数を集計したもの (ここでは遺伝子数の多かった上位10属を表示) 。分析対象であるChordodes属のハリガネムシは別属のハリガネムシ (Paragordius属、Gordius属) と相同性の高い遺伝子 (アミノ酸配列) を多く持つが、系統的に大きく離れているはずの宿主のカマキリがその次に多い。 (B) ハリガネムシの遺伝子 (DNA塩基配列) に対して、図で示した中で最も相同性が高い種を検索し、種ごとに該当するハリガネムシの遺伝子数を集計したもの。DNA塩基配列はアミノ酸配列に比べて変化しやすいため、分析対象であるChordodes属のハリガネムシは別属のハリガネムシ (Paragordius属、Gordius属) とであっても、大多数の遺伝子で有意な相同性が見いだせない。一方で、系統的に大きく離れているはずの宿主のカマキリでは、配列相同性のある遺伝子が多数見つかる。 この研究では、アミノ酸配列 (A) とDNA塩基配列 (B) の双方でカマキリと最も高い相同性が支持された1,342遺伝子をカマキリからの水平伝播候補の遺伝子とした。
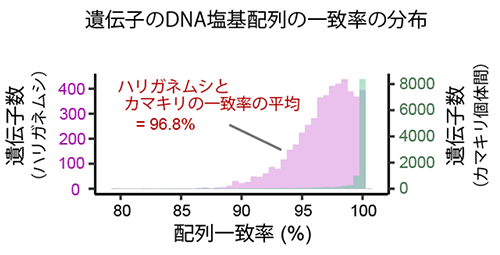
遺伝子 (DNA塩基配列) の一致率の分布を、ハリガネムシとカマキリ間 (マゼンタ) と、カマキリ個体間 (緑) で比較したもの。もしハリガネムシ分析試料にカマキリ組織が混入していれば、これらの試料から得たDNA配列はお互いに同じ類似度を示すと考えられる。しかしハリガネムシとカマキリではDNA塩基配列が少し異なる (平均の一致率96.8%) 。一方で、同種カマキリ個体間ではほとんどの遺伝子でDNA塩基配列が100%一致する。
この実験においてハリガネムシ分析試料にカマキリ組織が混入した可能性は、①宿主から脱出後、3日間の自由生活を経たハリガネムシから得たデータにおいても同様にカマキリの遺伝子との高い相同性が再現されたこと、②宿主個体間の遺伝子配列の一致度はほぼ100%であるのに対して、ハリガネムシの遺伝子は、宿主遺伝子との配列一致度が平均96.8%と、宿主と少しだけ異なっていたこと (図4) から、ほぼ排除できるものと考えられます。従ってこれらの結果は、ハリガネムシゲノムに宿主由来の遺伝子が組み込まれた「遺伝子水平伝播 (horizontal gene transfer, HGT) 」が進化の過程で生じていた可能性を強く示唆します。特に、ハリガネムシは系統的には遠い種であるにもかかわらず、他の昆虫よりもカマキリと高い相同性が見られたこと (図3B) と、②の通り同種間とは異なるものの近い配列一致度が得られたことからは、Chordodes属ハリガネムシの進化の過程において、遺伝子水平伝播が現在の宿主であるチョウセンカマキリからだけではなく、他種のカマキリからも起きた可能性が考えられます。
それでは、こうした水平伝播候補の遺伝子は、実際に宿主操作に関わっているのでしょうか。興味深いことに、水平伝播候補遺伝子は、候補以外の遺伝子 (カマキリ遺伝子と類似していない遺伝子) に比べて、宿主操作中に高い頻度で発現上昇していました (図5A)。またそれら遺伝子の発現量は、他の遺伝子の発現量に比べて高くなっていました (図5B)。こうした発現上昇が顕著な水平伝播候補遺伝子に宿主操作に関わるものがあるかを調べるため、既知の遺伝子の機能情報が集約されたデータベースでの検索を行いました。その結果、水平伝播候補遺伝子には、動物の行動活性に影響する「神経修飾物質[6] (生体アミン) 」、水面からの反射光への正の走性 (方向性のある運動) に関与すると指摘されている「光走性」、および行動操作の日周性に関与すると指摘されている「概日リズム」の機能に類別される遺伝子が含まれていました。すなわち、寄生虫のハリガネムシがこれらの水平伝播候補遺伝子を発現し、宿主の生体システムに介入させることで、宿主の入水行動が誘導されている可能性が考えられます。
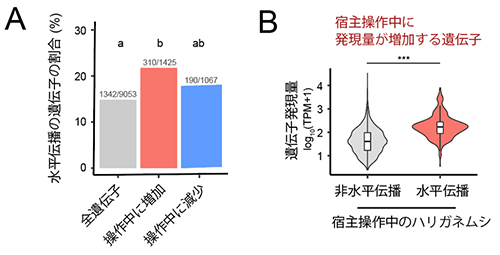
(A) ハリガネムシの全遺伝子 (グレー)、宿主行動操作中に遺伝子発現量が増加する遺伝子 (赤)、減少する遺伝子 (青) における水平伝播候補遺伝子の割合をそれぞれ示した。宿主行動操作中に発現量が増加する遺伝子には、全遺伝子に比べてより多くの割合で水平伝播候補の遺伝子が含まれる。 (B) 宿主操作中に発現量が増加する遺伝子における、水平伝播の可能性の有無に対応した遺伝子発現量の分布。縦軸はトランスクリプトーム解析で得られた相対的な遺伝子発現量、グラフの横幅はその発現量を示す遺伝子の頻度分布を表す。宿主操作中のハリガネムシで高発現する遺伝子は、水平伝播候補のものが多い。
注2) Biron, D., Marché, L., Ponton, F., Loxdale, H., Galéotti, N., Renault, L., Joly, C., and Thomas, F. (2005). Behavioural manipulation in a grasshopper harbouring hairworm: a proteomics approach. Proc. R. Soc. B 272, 2117-2126.
今後の期待
今回の研究から、ハリガネムシは、宿主からの大規模な遺伝子水平伝播によって獲得した遺伝子を利用して、宿主の生体システムに効果的に介入し、延長された表現型、つまり宿主の入水行動を発現している可能性が見えてきました。
遺伝子水平伝播は細菌などではごく一般的な現象として知られ、非病原性から病原性への進化や、抗生物質耐性の獲得などその多様性創出に寄与してきたと考えられています。一方で、多細胞生物間での遺伝子水平伝播は、トランスポゾンやウイルスを介した伝播の例などが増えつつあるもののまだまれな現象として認識されています。今回ハリガネムシで見いだされた大規模な遺伝子水平伝播は非常に驚くべき発見です。今後の研究により、水平伝播候補遺伝子がどのような仕組みでハリガネムシゲノムに取り込まれ、その後どのように進化してきたのかや、水平伝播候補遺伝子は宿主の行動改変に際してどのように機能しているのかが解明されれば、延長された表現型の分子機構とその進化の道筋が明らかになると期待されます。
補足説明
[1] 遺伝子水平伝播
生物の遺伝情報は通常は生殖を介して親から子へと伝えられるが (垂直伝播)、全く異なる個体間や他生物間で起こる遺伝子の取り込みのことを水平伝播と呼ぶ。
[2] 宿主操作
寄生生物が宿主の行動を改変する現象。本研究対象であるハリガネムシ-カマキリ以外に、トキソプラズマに感染したネズミはネコへの捕食回避行動が低下し、ネコに食べられやすくなる。これによってトキソプラズマの寄生先がネズミからネコに移動しやすくなると指摘されている例などが知られている。
[3] トランスクリプトーム
生体分子や細胞の挙動などの生体活動に関わる網羅的な情報をオミックスと呼び、トランスクリプトームは、オミックスのうち、一つのゲノム、または特定の細胞・組織・器官で発現する全ての遺伝子の転写産物 (RNA) を指す。
[4] 延長された表現型
ある生物個体の遺伝子が、その個体の表現型 (形態や行動など) の発現にとどまらず、他個体や周囲の環境の表現型発現に寄与すること。寄生生物による宿主操作では、寄生生物の持つ遺伝子が、宿主の形態・行動発現に寄与することと定義される。
[5] 分子擬態
病原菌や寄生虫などが産生する分子が、宿主が産生する分子と構造が類似していること。
[6] 神経修飾物質
神経細胞から分泌され、脳全体に持続的な効果を持つ神経伝達物質の総称。代表的なものにドーパミンやセロトニン、オクトパミンなどがあり、アミンと呼ばれる化学構造を持つ。
国際共同研究グループ
理化学研究所 生命機能科学研究センター 染色体分配研究チーム
- 基礎科学特別研究員 (研究当時) 三品達平 (ミシナ・タッペイ)
(現 客員研究員、九州大学大学院農学研究院 水産増殖学研究室 助教)
京都大学 生態学研究センター
- 准教授 佐藤拓哉 (サトウ・タクヤ)
国立台湾大学
- 助教 邱 名鍾 (Ming-Chung Chiu)
大阪医科薬科大学 医学部
- 講師 (研究当時) 橋口康之 (ハシグチ・ヤスユキ)
(現 准教授)
神戸大学 理学研究科
- 准教授 佐倉 緑 (サクラ・ミドリ)
- 学術研究員 岡田龍一 (オカダ・リュウイチ)
- 学部4年生 大石紗友美 (オオイシ・サユミ)
- 大学院生 佐々木淳成 (ササキ・アツナリ)
東京農業大学
農学部
- 教授 佐々木 剛 (ササキ・タケシ)
生物資源ゲノム解析センター
- 研究員 内山博允 (ウチヤマ・ヒロノブ)
福井県立大学 海洋生物資源学部
- 客員研究員 武島弘彦 (タケシマ・ヒロヒコ)
研究支援
本研究は、理化学研究所運営費交付金 (基礎科学特別研究員制度) で実施し、日本学術振興会 (JSPS) 科学研究費助成事業挑戦的研究 (萌芽) 「宿主操作の分子・神経メカニズムから解き明かす生態系のエネルギー流 (研究代表者:佐藤拓哉、19K22457)」、神戸大学国際共同研究強化事業「生物の集団動態を規定する鍵形質のゲノム基盤解明に関する国際共同研究」(研究代表者:佐藤拓哉)、科学技術振興機構 (JST) 創発的研究支援事業「寄生生物による生物機能創発機構の解明と制御への基盤研究 (研究代表者:佐藤拓哉、JPMJFR211C)」、東京農業大学生物資源ゲノム解析センター「生物資源ゲノム解析拠点 (研究代表者:佐藤拓哉)」などによる支援を受けて行われました。
論文情報
タイトル
DOI
10.1016/j.cub.2023.09.052
著者
Tappei Mishina*†, Ming-Chung Chiu*, Yasuyuki Hashiguchi†, Sayumi Oishi, Atsunari Sasaki, Ryuichi Okada, Hironobu Uchiyama, Takeshi Sasaki, Midori Sakura, Hirohiko Takeshima, Takuya Sato†
(*第一著者、†責任著者)掲載誌
Current Biology